 |
The Hissem-Montague Family  |
|
The Hissem-Montague Family |

A relative, Alan Hissem Jr., my 6th cousin, has completed the Big-Y test at FTDNA with a new terminal clade of G-FT411024. This is one step beyond the previous G-Z38868. My and Alan's Most Recent Common Ancestor (MRCA), Thomas Hesom, was born circa 1720 in Trenton, New Jersey. Our Y-DNA heritage is:
FT411024,Brian Fletcher, also G-Z38868, does not share in the latest clade. All members of the extended Hissem line in America (Hissam, Hessom, Hysham, Heysham, etc.) should share this latest Y-DNA type.
Z38868,
Z36217,
Z726,
CTS2230,
Z727,
Z1817,
CTS9737,
L497,
etc.
The Family Tree DNA site now claims that the Thomas Heesom (1743) family of East Yorkshire [hi Nicolette!] is also a member of the FT411024 clade per something referred to as 'Deep Clade-G.' Is this another test or simply an evaluation of markers discerned in the Heesom's 67 marker test (that haven't done the Big-Y)? I know that the Hissem family of America and the Heesom family of England share an MRCA, George Hesom, of Crofton, Yorkshire who was born circa 1600 (I have a good paper trail). Yet while FT411024 had branched off from Z38868 by 1500, according to FTDNA, they claim an MRCA date of 1800. Something is wrong here.
NEWS:A possible relative, Justin Rick Heysham of Melbourne, Australia, has said he will obtain a DNA test. Justin is one of the last four male Heyshams alive of the 'Merchants of Lancaster' line. If the Heesom/Line is related, we probably branched off during the 14th century's years of famine, plague and war.
The origin of the male line of our family, the Hissem Family, is a complex subject which covers not just hundreds or thousands of years, but hundreds of thousands of years. However, before we start this long and fairly torturous journey, I want first to identify the locus of the following discussion, which is northern England. This is, metaphorically, a stake in the ground for the earliest point of something like certainty in our story. It is from there that we can look forward, using written documents, to trace our family descent to the current day. It is also from there, looking backwards, that we can use biological and archaeological evidence to construct a coherent story of where the male line of our family came from and how they managed to get to the north of England.
On the previous page we claimed that our family name, Hissem, was a derivative of Heysham, which is a village in Lancashire, in northwestern England. The earliest recorded spelling of the village's name, in the Domesday Book of 1094, was Hessam. In the latter 17th century common variants included Heesom, Hesom and Heesham, all spellings used by our family during the same era.
If the above is valid, we can make an educated guess that our family lived in or near the village of Heysham in the 13th century. That is, they were there when surnames were first being developed. So, an ancestor, we'll call him John, became known, for administrative or legal purposes, as John of Heysham. This was done in order to differentiate him from John of, say, Halton, or John, William's son, or John, the tailor. See the Evolution of Names to learn more about this process. It is, however, difficult to say when the family first arrived in Heysham. Perhaps they had lived there for generations; perhaps they had just arrived. Looking at our DNA will help clarify the possibilities.
 DNA testing allows us to determine the degree of relationship between any two people, but more importantly it lets us trace the origins and development of man back tens of thousands of years and, combined with the archaelogy of ancient settlements, to follow his travels across the globe as well as his technological advancement.
DNA testing allows us to determine the degree of relationship between any two people, but more importantly it lets us trace the origins and development of man back tens of thousands of years and, combined with the archaelogy of ancient settlements, to follow his travels across the globe as well as his technological advancement.
Each one of us is a unique creation, a scramble of the genes provided by our mother and father. However, as we all remember from High School science, while our mother had two x-chromosomes, our father had an x- and a y-chromosome, the latter determining his sex. That means that a son can receive the y-chromosone that defines him as a man only from his father. The y-chromosome, then, identifies just the male line of descent, and does so back to the development of Y-DNA itself. This chromosome is passed from father to son relatively intact. However, over the generations mutations do occur, increasing in number with the centuries. Shared ancestry between two men can be determined based on measurements of their y-chromosome, and comparing the extent of their differences.
An individual's y-chromosome profile is known as his haplotype, from the Greek haplo, meaning simple or single. Individuals can be organized into groups of people sharing broadly similar haplotypes, called a haplogroup. Each haplogroup can trace their descent from a single man who originated the particular mutation that defines their group.
Using haplogroups it is possible to trace our family's migration from very early in man's evolution to the current era. To avoid my discussion of haplogroup evolution, below, jump to the Heesom/Hissem DNA type, G2a. Note that I don't have Y-DNA on my father-in-law, Lloyd Montague, but I expect that he is R1b of some flavor, as most Englishmen are.
By the way, the issue of what other function the Y-DNA may have, other than determining sex, has arisen again. A new study indicates that the Y-DNA may have some kind of control function over the operation of the other, autosomal DNA. See the auDNA page for more information about autosomal DNA.
| The Evolution of Man
The general scientific consensus today is that man evolved on the African continent, arising from an ancestor species of primate, not yet identified, whose descendents included both man and the great apes. This branching probably occurred some 5 to 7 million years ago. About 2 million years ago our hominin precursors, the Australopithecine, evolved into the homo genus. This genus produced a plethora of early men, including homo habilis, homo egaster and homo erectus, each iteration standing more erect, with a larger cranium and smaller jaw.  Anatomically modern humans, homo sapiens, or "wise man," emerged during the Middle Paleolithic, about 200,000 years ago. They then spread across the globe and by the late Paleolithic had replaced all of the archaic homo species, including the homo heidelbergensis, homo neanderthalensis and homo denisova. By the way, DNA analysis suggests that Neanderthal man, who inhabited Europe and parts of western Asia early in man's history, was not an ancestor of modern man, but a "dead-end" branch of the species that died out perhaps 50-60,000 years ago. Humans are the only living members of the genus homo. Recent research has indicated that modern man, as he migrated out of Africa into Europe and East Asia, encountered, and interbred with, archaic men, including Neanderthal, Denisovian, and homo heidelbergensis . . . as any woman will tell you, the sexual habits of men are simply scandalous. The "interchange" with Neanderthal apparenty happened about 60,000 years ago in the Middle East. One of the important traits gained was the tolerance for lactose in milk. An even more recent study indicates that, while modern Europeans do have Neanderthal antecedants, there is no evidence of Neanderthal in their Y-DNA. That is, either there were no male Neanderthal interbreedings, or all of their male progeny died out. Why did all other branches of the homo family die out? The modern answer is that humans killed them, or murdered them if you prefer the prejorative, but I think the salient point is that they all occupied the same ecological niche; success for one group entailed decline for the others. Archaic men disappeared because they could not successfully compete with modern humans. Human Differences That Made Our Civilization Possible
Modern humans are mammals of the primate order. That means they have stereoscopic vision and deft hands. However, they are differentiated from other primates and, most significantly, the Great Apes, by several factors.
|
It seems amazing to me, but we all share, far in the past, a single common male ancestor. Every person living today is a descendant of this one human being.
"The Y-chromosomal most recent common ancestor (Y-MRCA, informally known as Y-chromosomal Adam) is the most recent common ancestor (MRCA) from whom all currently living men are descended patrilineally. Y-chromosomal Adam is estimated to have lived roughly 236,000 years ago in Africa. By examining other bottlenecks most Eurasian men are descended from a man who lived 69,000 years ago. Other major bottlenecks occurred about 50,000 years ago and subsequently most Eurasian men can trace their ancestry back to a dozen ancestors who lived 50,000 years ago." - from WikipediaThe haplotype of this Y-chromosome Adam established the prime haplogroup, referred to as haplogroup A. For the purposes of this discussion I will pass over the more recently discovered pre-A groups, A0 and A00. He was not the "first man," but he does represent a choke point or bottleneck in our ancestry from which all men living today sprang.
| Single Nucleotide Polymorphisms
Single nucleotide polymorphisms, or SNPs, are the most common type of genetic variation among people. Each SNP represents a difference in a single nucleotide of the DNA. For example, a SNP may replace the nucleotide cytosine (C) with the nucleotide thymine (T) in a certain stretch of DNA. These changes are relatively rare. The SNPs of a man's Y-DNA identify his genealogical connections. Haplogroup classifications are based on SNPs. Short Tandem RepeatsShort Tandem Repeats, or STRs, is a spot on the DNA where a letter sequence is repeated, i.e. AGTAGT. STRs have a faster mutation rate than SNPs. For men have matching SNP results, their STR marker results determine how recently they are related. For this reason surname studies utilizes STRs. |
Our Adam lived during the Paleolithic era, or Early Stone Age. At this distant time the population of homo sapien males was very small, perhaps just a few hundreds, only one of whom had a line of descendents that has persisted to this day. I suspect that if we were able to find enough testable DNA material in the fossil record we would discover a plethora of potential early forebears whose lines were all doomed to perish at some point before today.
This should not, however, be confused with how man evolved.
"Evolution involves populations. The population slowly changes over generations. There is never a point where there is 1 male and 1 female of a new species within a generation. In fact, we can never pinpoint a generation where a new species started. For instance, at generation 1 there is species A and at generation 2,500 there is species B. But you cannot say 'here at generation 1,299 we have species A and at generation 1,300 we have species B'. Evolution never works like that." - from Paul Lucas, at Quora.comWhile evolution is a natural process, naming is done by humans and is arbitrary. We decide that at some point a certain accumulation of new traits decribes a unique type and so name it.
It should also be said that a single MRCA does not mean that this man descended from a single homogenous population. There is evidence that man evolved from several archaic homo populations, and in more than one location in Africa.
"When we look at the morphology of human bones over the last 300,000 years, we see a complex mix of archaic and modern features in different places and at different times," said Prof. Chris Stringer, researcher at the London Natural History Museum and co-author on the study. As with the material culture, we do see a continental-wide trend towards the modern human form, but different modern features appear in different places at different times, and some archaic features are present until remarkably recently.The resultant pre-modern peoples were able to interbreed, and most likely did so, perhaps with abandon (!). Our MRCA would have been the result of multiple interbreedings across these groups, combining the evolved characteristics of them all. By the way, the title of the article cited above is an attempt to blame modern humans for a 40,000 year trail of destruction from the loss of Ice Age mammals to the destruction of the rain forests of today. Part of their thesis is that we also murdered all the other homo species. I remember a book, or was it just a theory?, in my youth; I think it was called "The Killer Ape." It had a similar thesis. Ain't we awful? . . . or maybe we just out-competed the other "apes."
. . . To understand why human populations were so subdivided, and how these divisions changed through time, the researchers looked at the past climates and environments of Africa, which give a picture of shifting and often isolated habitable zones. Many of the most inhospitable regions in Africa today, such as the Sahara, were once wet and green, with interwoven networks of lakes and rivers, and abundant wildlife. Similarly, some tropical regions that are humid and green today were once arid. These shifting environments drove subdivisions within animal communities and numerous sub-Saharan species exhibit similar phylogenetic patterns in their distribution.
The shifting nature of these habitable zones means that human populations would have gone through many cycles of isolation - leading to local adaptation and the development of unique material culture and biological makeup - followed by genetic and cultural mixing." - from "Were other humans the first victims of the sixth mass extinction?" by Nick Longrich
The Paleolithic
The beginning of this era predates the emergence of modern man and marked the development of stone tool technology amongst the Australopithecine, a precursor to the hominids. It covers a period from about 3.3 millions years ago to the dawn of farming, 10-12,000 years ago. Prior to this time our earlier ancestors used tools made of the bones, teeth, and horns (or antlers) of their prey. This is called the Osteodontokeratic (i.e. Latin & Greek for bone-tooth-horn). Stone tools could be as simple as an unshaped rock or as complex as a flint axe. The first evidence of flaked rock tools were found with homo habilis remains dated to circa 2.1 to 1.5 millions years ago. Flint, or obsidian, is shaped by a process known as knapping. When struck by another rock, the flint fractures leaving a smooth surface. Repeated strikes can create an edge on the flint sharp enough to shave with. Other stones can be shaped as well, though with greater difficulty. Flint and obsidian became important commodities and were the foundation of the earliest trading networks. Also predating modern man was the creation and control of fire, which has been traced to from 700,000 to 120,000 years ago [that's a pretty wide range]. The bow and arrow, however, was a weapon of homo sapiens. It was developed about 65,000 years ago. Paleolithic Society Paleolithic society was based on the extended family, a structure required due to the demands of their young. A human child, unlike most animals, is helpless at birth and requires years of care before reaching maturity. It has been estimated that for a human infant to be born at the same level of development as a chimpanzee infant, the gestation period would have to be extended to 18 to 21 months. The family group developed to protect both the child and its mother during this vulnerable period. Even after weaning a child needs years of training and socialization, by mother, father, aunts and uncles, before it can become a full fledged member of the group. An individual tribe rarely had more than 30 members, many of whom would be children. Its leader would the most successful hunter and his chief advisors the eldest man and woman in the group, most likely his aged parents. Internal conflict was exclusively personal and could be handled one-on-one based on the good of the group. It was a communal society of a general equality in poverty. That is not to say that hierarchies did not exist. All animals have structures of dominance just as birds have their pecking orders. It is just that until larger numbers of people were grouped together the hierarchical order would of necessity be very flat. But human nature doesn't change. Strong men attempted to dominate those weaker than themselves and strong women, who today we'd call queen bees, tormented their frailer rivals. The religion of the Paleolithic was a hunter's cult of animal spirits who could be propitiated to make game available. Before a hunt a ritualistic dance would be held with the hunters wearing the pelts and antlers of their prey and mimicing the animal's movements. This action was either to 'call' the animal or to foretell, it was hoped, the results of the hunt. Cave art may have filled a religious purpose as well. Y-DNA haplogroups in Europe during the Paleolithic included BT, CT, C1, F and IJK. Haplogroup I is only found starting about 17,000 years ago. The Mesolithic This was a transition period between eras. In Europe it is usually considered to begin at the end of the last Ice Age, circa 10,000 BC. Haplogroups BT, CT, C1 and IJK had disappeared from Europe by this time while haplogroup I was undergoing a great diversification and expansion. R1a and R1b had emerged in Russia. The next period of history recognized is the Neolithic when agriculture was invented. |
 Our Adam was an anatomically modern human man, a homo sapiens, born quite late in the history of the Earth. He differed from archaic humans by having a higher forehead and larger cranial capacity, a smaller face, and more delicate skeleton. He stood from 5 feet 4 inches to 6 feet tall.
Our Adam was an anatomically modern human man, a homo sapiens, born quite late in the history of the Earth. He differed from archaic humans by having a higher forehead and larger cranial capacity, a smaller face, and more delicate skeleton. He stood from 5 feet 4 inches to 6 feet tall.
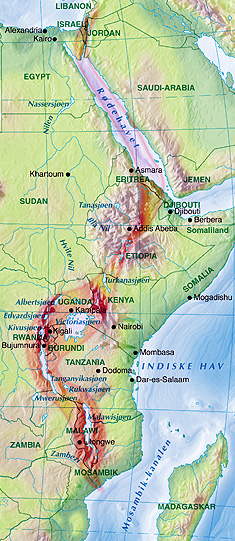
The human population of which Adam was a member first arose in east Africa. This is the region of the Great Rift Valley, pictured to the right, in present-day Ethiopia, Kenya, & Tanzania, where some of the earliest hominid remains have been found. Think of Dr. Leakey's discovery of an Australopithecus nicknamed "Lucy." The latter species was the precursor of homo habilis.
From the initial haplogroup, type A, there evolved the rest of the twenty male human haplogroups now existing, lettered from B through T. That is,
"Each haplogroup originates from, and remains part of, a preceding single haplogroup (or paragroup). As such, any related group of haplogroups may be precisely modelled as a nested hierarchy, in which each set (haplogroup) is also a subset of a single broader set." - from WikipediaSo, haplogroup B is a subset of A, just as haplogroup C, as we'll see, is a subset of B. Refinements within a haplogroup are represented by additional numeral and letter combinations, that is, for example, A > A1 > A1a > A1a1. Each of these subgroups has a single male at its origin whose mutation defines the subgroup. As more subtypes have been identified this system has proved clumsy; a type designation of twelve or more alphanumerics is difficult to read or remember. Increasingly types are defined by a shorthand method of the group, i.e. A, and the mutation that defines the subtype, M171 for instance, displayed as A-M171. How it is decided that a particular mutation merits the creation of a new haplogroup vice another subgroup of the current haplogroup I could not say.
There is, of course, also a most recent common female ancestor, a Most Recent Common Ancestor-Eve, who is defined based on the matrilineal transmission of mitochondrial DNA. This type of DNA, inherited only from the mother, is passed down to both male and female children. However, I haven't yet studied that concept adequately to understand it nor to use it for our family. See the webpage The mtDNA Story for what I know on the subject.
Of course, identifying a direct-male and direct-female line of ancestry through our Y-DNA and mitochondrial DNA leaves out a large number of our ancestors. At just the level of our grandparents it ignores the descent of our paternal grandmother and maternal grandfather. A way to capture these other influences from our past is through the use of autosomal DNA. These are the twenty two pairs of non-sex chromosomes that are inherited from both parents. An analysis of these will show what percentage of your ancestry is, for instance, Western European or Native American. See The Autosomal DNA Story for a discussion of this.
From Africa modern man migrated to settle the rest of the planet; haplogroup A, found only in Africa, pre-dates this "out of Africa" migration.
| Out of Africa Theory Modified
Recent archaelogical exploration has unearthed the remains of anatomically modern humans in Arabia and the Levant that are much older than the presumed out of Africa movement. Those in Arabia were dated to greater than 85,000 years ago and in the Levant to greater than 175,000 years ago. This has been called a failed dispersal because no descendants have been found for these early migrators. |
Haplogroup BT derived from a mutation to A about 70,000 years ago. It is the common ancestral type of all early migration out of Africa. The basal paragroup BT has not been found in modern populations, the T in the name denoting this fact. There is, then, a 130,000 year gap from the initial haplogroup to its next major subgroup. As we'll see, subsequent haplogroups developed far more rapidly. What stifled the development of mutations to the Y-DNA at this early time or is this simply the result of the low population?
| Genetic Chokepoints and Catastrophe Theory
At some point, and it's apparently around 70,000 years ago, the human population faced a sharp reduction in its size. "Unlike our close genetic relatives - chimps - all humans have virtually identical DNA. In fact, one group of chimps can have more genetic diversity than all of the six billion humans alive today. It is thought we spilt from a common ancestor with chimps 5-6 million years ago, more than enough time for substantial genetic differences to develop. The absence of those differences suggests to some researchers that the human gene pool was reduced to a small size in the recent past, thereby wiping out genetic variation between current populations." - from BBC New Online Science Editor One possible explanation for this bottleneck in human development may be the eruption of a supervolcano at Lake Toba in Sumatra approximately 70,000 years ago. The resultant cooling of the planet, a volcanic winter, may have caused a large die-off and restriction of the human population to a few tropical refugia. A drastic reduction in the population, to perhaps as few as 40 breeding pairs, resulted in an equally drastic reduction in genetic diversity. It was not until the late Stone Age that the human population recovered to its pre-catastrophe levels. The Cheetah Chokepoint This isn't necessarily relevant to our discussion, but I think it interesting that the cheetah, the super-fast feline of the African savanna, also experienced a near-extinction event. About 10,000 years ago, when many large mammals were dying out as the planet warmed, the cheetah population crashed. Today they suffer from extremely low genetic diversity and resultant deformities. |
Haplogroup B and CT derived from BT. CT contains a mutation present in all haplogroups, except A, and B. It developed in East Africa about 60,000 years ago.
"No male in paragroup CT has yet been discovered, which means in other words that all men in this haplogroup are also defined as being in one of the several major branch clades. All known surviving descendant lineages of CT are in one of two major sub-clades, CF and DE." - from Wikipedia
| The Life of Early Man
The earliest men were hunter-gatherers, that is, they hunted game and gathered roots and berries for their subsistence. They were a mobile people, tracking the ever moving herds and staying in one place only until the food that could be gathered in the area had been exhausted. Being mobile they had few possessions; some skins for clothing, a few tools or weapons made of bone and antler, and later of stone, wooden spears with fire-hardened tips, some baskets woven of reeds to carry food, a gourd to carry water. They had no shelter, unless it was a rapidly built lean-to made of leaves and branches or a hole in the ground covered with similar materials. The band or tribe was probably a small one, numbering no more than 30 individuals. There would be several adult males and a similar number of adult females, and a variety of adolescents, small children, and infants. If the group grew any larger it would tend to splinter. Most of the food was gathered by the women from the local brush. Accompanied by their children, they spent most of the day digging for roots and tubers, and gathering whatever fruit or nuts could be pulled from the trees. The men would leave on long hunts. Lacking effective weapons they had to track the animals they wounded for many miles, and often for many days, until the animal, exhausted, could be cornered and killed. Man, while a relatively poor sprinter, is an unparelleled distance runner and can, given time, out run a gazelle. The meat, fat and marrow they brought home was a welcome respite to the usual vegetable fare. The band, lacking means to preserve and store the meat, would gorge. As often as not though, the men came back empty handed. Humans had to compete not only with the other predators, such as lions, bears and wolves, but with other early hominids like the Neanderthal. For the majority of man's time on earth we have been hunter-gatherers, existing at a very simple level. While this life could be hard, it also had its advantages. People only worked when they needed food, the rest of the time they might laze about, much like a pride of lions that has had a recent kill. Note that in later periods people considered hunting to be a recreation, a relief from the stress of work and responsibility. The change to this mode of living happened relatively recently, as will be discussed below. By the way, the dog was the first animal domesticated. This happened 15,000-35,000 years ago, depending on your source, during this hunter-gatherer phase. The human-dog combination made for an unbeatable hunting team, combining human ingenuity and wolf speed and ferocity. Watch one of those police video shows ["Bad boys, bad boys"] and note the perp's response when the police release the dog.  Human groups had to meet and compete with other humans. Like other territorial animals, conflicts would occur between other roving bands of humans as they sought to maintain or expand their hunting domain. As has been seen in research with chimpanzees, animals other than man do make war and plan the conquest of competing groups. Humans fought, perhaps frequently, though the primitive nature of their weapons meant these battles were short. However, the loss of hunting territory could mean eventual death for the entire group. There must have been some friendly coordination between groups, of course, if only to trade prospective sexual partners to prevent inbreeding. A patrilocal society is one in which women move to the tribe of their husband. A matrilocal society is one where men move to the tribe of their wife. |
The move out of Africa may have been due to a fluctuation in climate. About this time the most recent glacial period ebbed in force, resulting in a warmer/wetter environment. The Sahara bloomed into grasslands, drawing grazing animals north, who were followed by their predators, including man. The exact route they followed remains unknown, but our ancestors eventually followed the expanding grasslands and plentiful game into the Middle East.
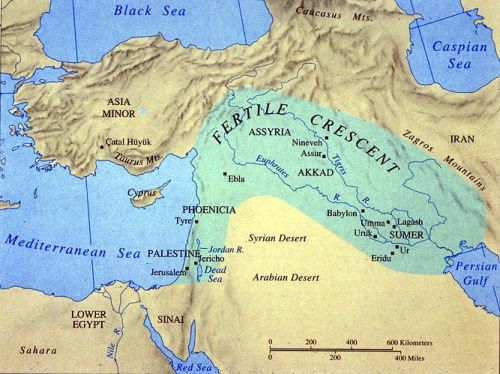
The Great Rift Valley runs up through East Africa to the mouth of the Red Sea, at Djibouti. Across that sea is the Arabian penisula. During this period was this a "dry-shod" route? Or did our ancestors move up the Nile Valley, and then across the Sinai? The latest theory claims that the first, and perhaps only, successful pulse of humanity out of Africa, haplogroups CT, DE and CF, drove straight east, along the northern edge of the Indian Ocean, to Southeast Asia. It was only later that some humans turned around and migrated back west into the region of the Fertile Crescent.
Haplogroups C, D, E and F evolved from CT. Haplogroup F first appeared in North Africa, the Levant and the Arabian penisula about 50,000 years ago. Not long after the emergence of F another cold and arid period intervened. Drought hit Africa and the grasslands reverted to desert, effectively closing the gateway into the Middle East.
The Lake Toba volcanic eruption, which occurred about 70,000 years ago, and a subsequent population bottleneck, has been conjectured as an impetus to the break up of haplogroup CT, via CDEF, into C, DE and F.
| Genetic Chokepoints, continued
A genetic chokepoint results in a smaller population in which mutations, when they occur, get passed through a very large percentage of its members. This can result in fairly fast shifts in the population, especially if a few dominant males are responsible for the majority of the offspring. Anthropologists have proposed that such bottlenecks were responsible for the rapid development of hominids. The action of a chokepoint may result in what is known as the 'founder effect.' This occurs when a small, isolated population drastically diverges from the original population. As humans spread across the planet, scientists believe that our population experienced multiple bottlenecks and, as a result, a serial-founder effect kicked in to create the diversity we currently see in the human race today. One bottleneck occurred when a small group of humans left Africa. Another happened when this group split up in the Middle East, with one group heading to Europe and the other to Asia. Others occurred when humans left Southeast Asia for Austronesia, crossed the Beringia land bridge into Alaska, and spread into South America through what is now Panama. "A 2005 study from Rutgers University theorized that the pre-1492 native population of the Americas are the descendants of only 70 individuals who crossed the land bridge between Asia and North America." - from WikipediaBecause of this, African populations tend to have far more genetic diversity in their DNA than populations native to the Americas. |
Over the next 20-30,000 years haplogroup descendants of haplogroup F arose in the Near East, history's Fertile Crescent, then fanned out to populate the globe. These include all members of Haplogroups G through T, or 90% of the world's population.
Some haplogroups dominate their regions of settlement, especially in those instances where subsequent migration by other haplogroups is cut off. Haplogroup Q, for instance, dominates in the native population of the Americas. Haplogroup N does the same in the extreme northern latitudes of Eurasia, from Finland to Siberia, while haplogroups S and M are found predominantly in New Guinea and the nearby islands. Europe is dominated by just three haplogroups, I, R1a and R1b, that define northern Europe & the Balkans, eastern Europe and western Europe, respectively.
Some haplogroups are quite large, like R1 which spreads across Eurasia or O which dominates in China and Southeast Asia, while others are rather rare, like F, despite its having founded so many other haplogroups, and P, which was the parent of the giant Q and R groups.
Phylogenetic Tree of Human Y Chromosome DNA Haplogroups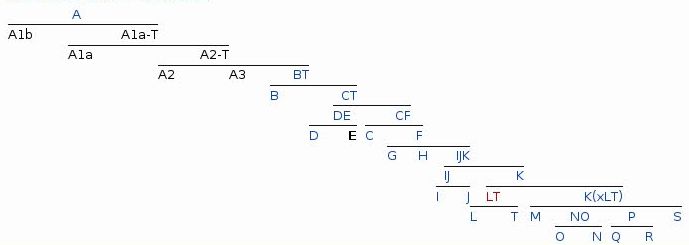
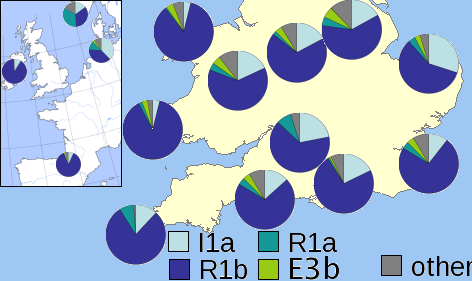 Since I've claimed on the previous page that the Hissem family emigrated to America from England, it's important to know what the Y-DNA break-out is for that country. At right is a map showing the distribution of Y-DNA in modern England and Wales.
Since I've claimed on the previous page that the Hissem family emigrated to America from England, it's important to know what the Y-DNA break-out is for that country. At right is a map showing the distribution of Y-DNA in modern England and Wales.
Haplogroup R1b is today the most common type in England, as it is in most of Western Europe.
Haplogroup I1a is the next most frequently found. I1 is the signature type in Sweden, Norway, Denmark and northern Germany, and is evidence of the Anglo-Saxon, Viking, and Norman invasions of the historic era. Note however that in only one region on the map at the right does haplogroup I arise above 25%. This region is known as East Anglia, for the German Angles who settled it.
- I had been taught in school that the Anglo-Saxon invasion replaced the native Celtic population, with the defeated remnants being forced into refuges in Wales, Scotland, and Cornwall. Apparently the majority of the natives actually remained in place while the invasion swept past them.
R1a, the major haplogroup of Eastern Europe, is far less common in England. The small amount seen may be due to modern migrations or it might have been brought by Viking populations who also incorporate this type.
E3b, the fourth most often found, is the only E-type haplogroup found in Europe in significant numbers. It came out of the Near East and North Africa, and might owe its presence in England to military and commercial populations brought in during the Roman occupation of the 1st through 4th centuries AD.
Haplogroup Percentages in England:The Heesom family of England and the Hissem family of America are part of haplogroup G. See Y-DNA Haplogroup G and its Subclades for the full haplogroup tree.
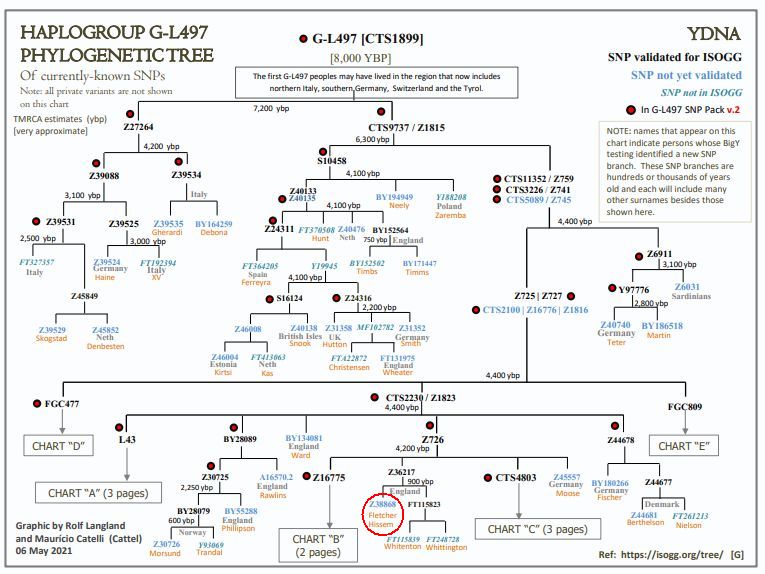
Members of the family who were tested are Crispin Heesom, of the East Yorkshire Heesom Family, Alan Hissem, of the Thomas Hissam of Thomas Family, James Olin Hissom, of the David Hissom of David Family, and myself, Steve Hissem, of the Jesse Hissam of David family. We are part of the G-Z726 subclade, under the umbrella of G-L497, which originated in the northwestern corner of continental Europe in about 2,600 BC. More significantly, we are part of G-Z36217, the immediate descendant of G-Z726, which evolved in about 2,000 BC and which is, so far, exclusively English. For this reason I believe our family migrated to Britain between 2,600 and 2,000 BC. For a more extensive discussion of this issue see The Bell Beaker Culture 2,750 - 1,900 BC. Finally, the terminal clade identified for the family is G-FT411024. It evolved in about 1500 AD. FTDNA indicates an MRCA of 1800 AD, but I have a paper trail that shows an MRCA of 1600 AD.
The latest G2a-L497 chart from Family Tree DNA:
Our ancestors were not part of the original, Paleolithic settlement of England, nor do they make up a significant proportion of England's population today.
So, where did our father-line ancestors come from originally?
Our family type, haplogroup G, is defined by a mutation at the M201 site on the chromosome and is a widespread, but relatively uncommon type. Members of the haplogroup share a common ancestor who developed a mutation from the earlier haplogroup F about 45,000 years ago [actually it evolved via an intermediary haplogroup, GHIJK, but that just confuses things]. This was during the Pleistocene, a glacial period of increasing cold of Arctic or Tundra conditions in Europe, but before the bitterest cold of the Last Glacial Maximum, which lasted from 25,000 to 17,000 BC. The latter event pushed any human population that existed in Europe south, to refugia at the far edges of the continent.
By the time our haplogroup emerged our male-line ancestors had already left Africa and made their way north into today's Middle East, a relatively rich region, able to support a large population.
| The Upper Paleolithic
The period from 50,000 to 10,000 years ago has been called the Late Stone Age or Uppler Paleolithic, and pre-dates the discovery of agriculture. It was in this period that anthropoligists believe mankind first began to demonstrate the ability to use complex symbolic thought and express cultural creativity, seen in cave paintings and the carving of voluptuous fertility figures.
 Man's tools became more diversified at this time, with specific tools for specific purposes, including the first use of fish hooks. This improved tool technology resulted in more efficient hunting, a better food supply, and a population explosion which probably doomed any remaining archaic men. |
Exactly where in the Middle East haplogroup G may have originally emerged has been widely and fiercely debated. One group argues that it was somewhere in the region of the Caucasus mountains. This is the area of the greatest concentraion of G haplotypes today. Haplogroup G is most common in North Ossetia and specifically, the town of Digora, with an average frequency of 74%. 29% of the Kabardinian and Balkarian peoples of the northwestern Caucasus are G types, and Azerbaijan and Armenia also have high concentrations of the haplogroup.
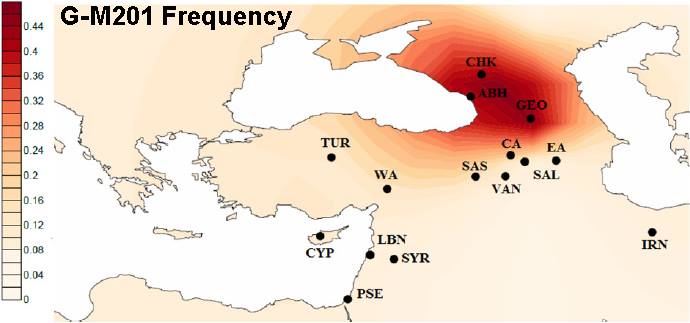
The other camp argues that it is a mistake to take this concentration of modern haplogroup G samples as proof of origin. They hold that the haplogroup arose, or at least had its population break-out, in the Levant of modern day Syria, Lebanon and Israel.
"Notice that while G-M201 reaches a high frequency (of about 50%) only in some areas of the Western and Central Caucasus, the variance of G for 8 (?!) STRs is highest in the Southern Levant." - from the Facebook G-M201 webpage
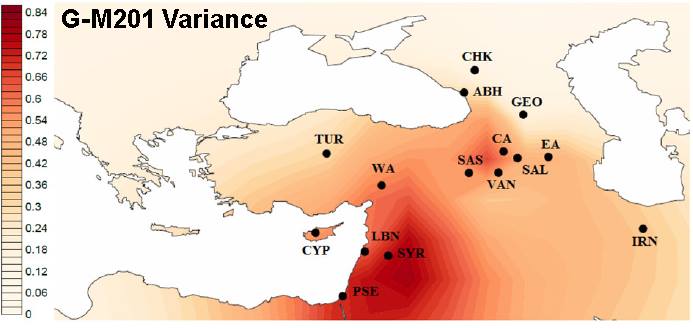
Unfortunately, not everyone seems to use the same data. At the eupedia.com website a highly regarded researcher, Maciamo Hay, says that,
"The highest genetic diversity within haplogroup G is found in the northern part of the Fertile Crescent, between the Levant and the Caucasus, which is a good indicator of its region of origin." - from eupedia.comSo, in sum, haplogroup G's origin was in the north or the south or in-between . . .
Up until the last few years such theories were based on the use of DNA extracted from modern subjects and making assumptions based on current population densities. New techniques have since been developed that allow the use of DNA from ancient burial sites and these show little correlation with current dispersal patterns. One of the DNA field's main researchers has said,
". . . the homeland of this haplogroup [G] has been estimated to be somewhere nearby eastern Anatolia, Armenia or western Iran, the only areas characterized by the co-presence of deep basal branches as well as the occurrence of high sub-haplogroup diversity." - from "Distinguishing the Co-Ancestries of Haplogroup G Y-chromosomes in the Populations of Europe and the Caucasus," 2012, by Rootsi, et alSo, perhaps we can agree to say that the locus of haplogroup G's emergence was somewhere in the Fertile Crescent of the Middle East, but most likely at its extreme northern tip in south-eastern Turkey, or Anatolia.
| Anatolia
The portion of Turkey to the east of Anatolia is generaly referred to as East Anatolia. |
Haplogroup G had a slow start, evolving in isolation for tens of thousands of years, with a small population. A smaller population meant fewer total mutations and a more homogeneous group.
"As of late 2016, there were 303 mutations defining haplogroup G, confirming that this paternal lineage experienced a severe bottleneck before splitting into haplogroups G1 and G2." - from eupedia.comAs of 2021 there are 317 mutations, or variants, defining the basal haplogroup G. A severe bottleneck simply means that few male lines were developed from the original source. It may be chance, scarcity, or adverse events that resulted in so many male descendents failing to produce more than the minimum number of male heirs for so long.
Further on this subject, haplogroup G's bottleneck can be best illustrated using G-M201's branch length. Branch length is a measure of divergence or genetic change in a lineage. The units of branch length are the number of variants or mutations. The longer the length of the branch, the more genetic change has occurred without sub-clades forming. M201's branch is 317 variants long which equates to about 23,000 years between its emergence and that of its first subclade. See more about branch length below.
Our earliest G-type ancestors would still have been Stone Age Hunter-Gatherers, organized into small bands of semi-nomadic people living off the land. Genetic researchers refer to them as Anatolian Hunter-Gatherers (AHG). That group makes up one of the three ancestral types to populate Europe. The others were Western Hunter Gatherers (WHG) who entered Europe during the Paleolithic and Ancient North Eurasians (ANE) who entered in the early Bronze Age.
Before talking about how haplogroup G spread and how we got into Europe, it is important to remember that at this early period the geography was different. The mountain passes of the Caucasus were obstructed by fields of ice, while a lower sea level had created a land bridge between Asia and Europe at the Bosphorus. The Black and Caspian Seas were larger than today, despite the seas being lower, because at this time they were fresh water lakes. Later, both diminished as the climate heated, then, circa 6,500 BC, that is, in the midst of the Middle Eastern Neolithic, a rising sea, fed by melting glaciers, tipped over the edge of the Bosphoros and turned the Black Sea salty.
| The Earth's Climatic History
A warming of the Earth, and subsequent glacial retreat, began in about 12,000 BC. The warming was shortly interrupted by a sudden cooling, known as the Younger-Dryas. Evidence from ice cores in Greenland show that about 12,000 to 13,000 years ago, something drastically disrupted our climate. The change was sudden, and it wiped out the remaining great fauna on Earth at the time, including the wooly mammoth, the saber-tooth tiger, and the giant sloth. Scientists speculate that this cooling may have been caused by the release of fresh water trapped behind ice on North America into the North Atlantic Ocean. The release altered vertical currents in the ocean which exchange heat energy with the atmosphere. A later theory was that the cooling was triggered by a comet strike. Fragments of a large, disintegrating asteroid or comet struck North America, South America, Europe, and western Asia around 12,800 years ago. Multiple airbursts/impacts produced the Younger Dryas boundary layer, depositing peak concentrations of platinum, high-temperature spherules, meltglass, and nanodiamonds, forming an isochronous datum at more than 50 sites across about 50 million km2 of Earth’s surface. Some scientists have proposed that this event triggered extensive biomass burning, a brief impact winter and the Younger Dryas abrupt climate change, contributed to extinctions of late Pleistocene megafauna. The warming resumed by 8,500 BC and between 5,000 and 3,000 BC average global temperatures reached their maximum level during the Holocene and were 1 to 2 degrees Celsius warmer than they are today. Climatologists call this period the Climatic Optimum. During the Climatic Optimum, many of the Earth's great ancient civilizations began and flourished. In Africa, the Nile River had three times its present volume, indicating a much larger tropical region. From 3,000 to 2,000 BC a cooling trend occurred. This cooling caused large drops in sea level and the emergence of many islands and coastal areas that are still above sea level today. A short warming trend took place from 2,000 to 1,500 BC, followed once again by colder conditions. Colder temperatures from 1,500-750 BC caused renewed ice growth in continental glaciers and alpine glaciers, and a sea level drop of between 2 to 3 meters below present day levels. The period from 750-150BC saw warming, however temperatures did not get as warm as the Climatic Optimum. During the time of the Roman Empire, 150 BC-300 AD, a cooling began that lasted until about 900 AD. At its depth, the cooling caused the Nile River and the Black Sea to freeze, 829 and 800-801 AD respectively. The period 900-1200 AD has been called the Little Climatic Optimum. It represents the warmest climate since the Climatic Optimum. During this period, the Vikings established settlements on Greenland and Iceland. The snow line in the Rocky Mountains was about 370 meters above current levels. A period of cool and more extreme weather followed the Little Climatic Optimum. A great drought in the American southwest occurred between 1276 and 1299. There are records of floods, great droughts and extreme seasonal climate fluctuations up to the 1400s. From 1550 to 1850 AD global temperatures were at their coldest since the beginning of the Holocene. Scientists call this period the Little Ice Age. During the Little Ice Age, the average annual temperature of the Northern Hemisphere was about 1.0 degree Celsius lower than today. During the period 1580 to 1600, the western United States experienced one of its longest and most severe droughts in the last 500 years. Cold weather in Iceland from 1753 and 1759 caused 25% of the population to die from crop failure and famine. Newspapers in New England were calling 1816 the year without a summer. The period 1850 to present is one of general warming. - from PhysicalGeography.net |
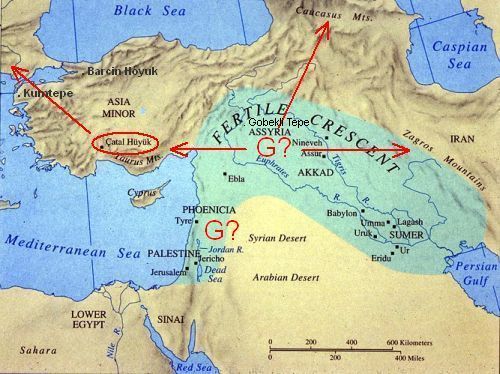
The G-type parent clade, G-M201, finally branched into G1 and G2 types about 23,000 years ago. The population holding the G1 mutation would migrate east, out of the Fertile Crescent, into Iran, crossing over the Zagros mountains. Later they also moved north into Kazakhstan and further east into western India.
Haplogroup G2, a mutation at P287 [L89], would move west into Anatolia and Cyprus, and northeast into the Caucasus Mountains. As you'll read, below, our ancestors were part of the group that moved west.
However, it was the descendants of these two main branches that would migrate and none of these movements would begin until the greatest invention in man's history had been made, that of agriculture. This inspiration ushered in the era of the Neolithic, or New Stone Age, but it wouldn't begin until another 10,000 years had passed.
| The Neolithic
The invention of farming and establishment of settled communities brought problems as well as advantages. With a guaranteed source of food the population boomed, but as it became dependent on a limited variety of foods, the general health was, paradoxically, poorer than that found amongst the hunter-gatherers. With an increasing population the margin of error between a good harvest and famine also narrowed dramatically. Finally, farming is hard, back-breaking work, requiring continuous vigilance against bad weather and pests. In future times people might hunt simply for the fun of it, but if the poor could escape the farm by fleeing to the cities, they did so. Today people garden as a hobby because they don't have to depend on it (I'm gardener and I know). It has been said that the Neolithic revolution was a trap. Once a society made the transition to farming they could never go back to hunting and gathering; their populations had grown too great to be supported by the old methods. Villages too had their disadvantages. They were targets for predatory nomads who could smash and grab before the farmers could organize to resist them. The history of Europe is of slow economic growth, periodically swept away by riders out of the east, or seaborne raiders from the north. It was only after about 1350 AD that Europeans were able to put an end to nomadic raids. Neolithic Society The change to farming and permanent settlements resulted in political change as well. A Paleolithic society would tend to be a communal one of general equality in poverty. The Neolithic unleashed a surplus not only of food, but of the crafts that there was now time to develop. Trade resulted, which increased the wealth. However, over time wealth stratified the society into the successful and unsuccessful. The former became the leaders which accelerated their rise in wealth. While a hunter-gatherer band was a family group, the farmers' world was one of many families and competing interests. As the size of a Neolithic community increased so did its complexity. Intermediaries had to exist to resolve inter-family, or inter-clan disputes. Others were needed to resolve arguments between farmers, artisans and merchants. And, because family ties were still the most important, once a man got a position of power he would try to keep it in his family group for succeeding generations. It was not long, therefore, before hereditary leaders arose. The Neolithic revolution had a sexual aspect as well. In the period of population growth that followed the spread of agriculture, circa 8,000-4,000 BC, a bottleneck developed in the male line resulting in modern populations having many more female ancestors than male of this era (on the order of 17 to 1). Researchers believe that this drastic drop in male diversity was caused by the rise of an elite few males who accumulated wealth, power and, subsequently, access to females, at the expense of other males. The elite then passed on their wealth, and reproductive success, to successive generations of sons. Apparently our fathers were amongst this lucky elite since we're here now. A stellar example of this elite advantage is evidence that Genghis Khan's DNA is present in about 16,000,000 men alive today. There are another 10 men who similarly dominate the genetic pool, though their identities are unknown. Male diversity recovered, though even in more recent history a global average of 4 women have reproduced for every 1 man. At this time religion too grew from a hunter's cult of animal spirits into one of hierarchial gods who governed the prospects for a good harvest. Farmers are the original pessimists of the world so it is not surprising that their gods were unpredictable and mean spirited, kinda like the weather. The Greek myths are replete with stories of gods that act like spoiled children or your most regrettable relatives. Y-DNA haplogroups in Europe during the Neolithic included G2a, as well as the hunter-gatherer groups already present during the Mesolithic. The next period of history recognized is the Chalcolithic, staring in about 4,500 BC, when copper metallurgy was developed. |
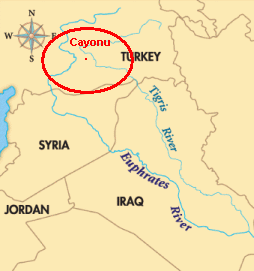 We've already said that our haplogroup probably arose at the northern end of the Ferile Cresecent. That sounds like the headwaters of the Euphrates and Tigris rivers on or near the Armenian plateau in eastern Anatolia. The ancient city of Cayonu Tepesi, it dates to before 10,000 BC, is located there; see the map to the left. The people living in Cayonu are believed to be the first farmers of Anatolia.
We've already said that our haplogroup probably arose at the northern end of the Ferile Cresecent. That sounds like the headwaters of the Euphrates and Tigris rivers on or near the Armenian plateau in eastern Anatolia. The ancient city of Cayonu Tepesi, it dates to before 10,000 BC, is located there; see the map to the left. The people living in Cayonu are believed to be the first farmers of Anatolia.
As far as I know, no usable DNA has been recovered from this site.
| Cayonu Tepesi
Genetic studies of emmer wheat, the precursor of most current wheat species, show that the slopes of Mount Karaca, which is located in close vicinity to Cayonu, was the location of first domestication. This was also possibly the place where the pig was first domesticated. An analysis of blood found at the site suggested that human sacrifice occurred there (!). |
Haplogroup G2a, which is an ancestor to our family subgroup, is a mutation at P15. It emerged about 18,000 years ago and is now believed to be a marker for early Neolithic farmers in Europe.
"There has so far been ancient Y-DNA analysis from Early Neolithic Anatolia, Iran, Israel, Jordan as well as most Neolithic cultures in Europe (Thessalian Neolithic in Greece, Starcevo culture in Hungary/Croatia, LBK culture in Germany, Remedello in Italy, and Cardium Pottery in south-west France and Spain) and all sites yielded a majority of G2a individuals . . . This strongly suggests that farming was disseminated by members of haplogroup G at least from Anatolia/Iran to Europe." - from Haplogroup G2aThis was more firmly stated recently,
"The farmers of Europe appear to be a very closely related group, that derives from a potentially singular source." - from "European Farmers Part I; Mediterranean vs Danubian" in Population GenomicsThat singular source being the G2a farmers of Anatolia. Neolithic settlements identified in Anatolia include Catalhoyuk, Cayonu Tepesi, Nevali Cori, Asikli hoyuk, Boncuklu hoyuk, Hacilar, Gobekli Tepe, Norsuntepe, Kosk, and Mersin. Catalhoyuk, in Central Turkey, is considered the most advanced of these, and Cayonu Tepesi in the east, at the northern end of the Fertile Crescent, the oldest. Hacilar, in Western Turkey, followed Cayonu Tepesi and has been dated to 7,040 BC. - based on Wikipedia.
These Anatolian farmers were the descendants of the Anatolan Hunter-Gatherers (AHG) mentioned earlier.
The Neolithic Farming Revolution
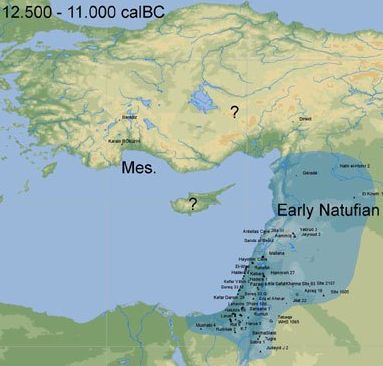
In the period between the Paleolithic and Neolithic eras, a population in the Levant, dubbed the Natufian culture, were able to give up their migratory habits and settle into a sedentary lifestyle. They were able to do so, despite continuing as hunter-gatherers, because the Middle East was at this time, unlike today, cool and lush with vegetation and animal life. The people hunted the numerous game and gathered the grains of the many wild cereal plants that grew there without the need to move on as the vegetation was depleted. In this friendly environment they forsook their previously nomadic lifestyle, settled down, and multiplied quickly.
By the way, there's some conjecture that it was women who invented farming. This makes sense since in most hunter-gatherer populations the men go out hunting game and women, encumbered by their children, seek roots, berries, and cereals in the nearby brush. As such it was more likely that it was they who cleared away the scrub that was crowding out the food plants they had been harvesting. From DNA testing of the sparse remains available, the Natufians of the Levant appear to have been haplogroup E1b, so they are not ancestors of our family. |
Who were the ancestors of these Anatolian farmers?
"The first farmers of the southern Levant (Israel and Jordan) and Zagros Mountains (Iran) were strongly genetically differentiated, and each descended from local hunter-gatherers . . . Among first farmers, those of the Levant trace ~2/3 of their ancestry to people related to Natufian hunter-gatherers and ~1/3 to people related to Anatolian farmers . . ." - from "The Genetic Structure of the World's First Farmers" by Lazaridis et al 2016So, while there had been little mixing of farmer populations at the extreme eastern and western ends of the Fertile Crescent, those to its north, who later found their way into Anatolia, were a mixture of both and of local hunter-gatherers. Trade was probably the driver of this intercourse. Ancestry here refers to autosomal DNA, the non-sex chromosomes, which helps identify all ancestors, male, female, and those not on the direct line of descent. This is what is used for those genetics tests that show you're 10% American Indian, for example.
"When it came to actually looking at the ancestral breakdown of Anatolians, Lazaridis et al. (2016) came up with a very solid model where Anatolians were a mix of lineages related to Ganj Dareh [Zagros mountains of Iran], Levant Neolithic, and WHG [Western Hunter Gatherer], with mixture proportions of 0.387, 0.339, and 0.274, respectively." - from "The First Farmers, with a Focus on Anatolia" at populationgenomics.blog
On the other hand, I find this,
"We show that Western Iran was inhabited by a population genetically most similar to hunter-gatherers from the Caucasus, but distinct from the Neolithic Anatolian people who later brought food production into Europe. The inhabitants of Ganj Dareh made little direct genetic contribution to modern European populations, suggesting those of the Central Zagros were somewhat isolated from other populations of the Fertile Crescent. Runs of homozygosity are of a similar length to those from Neolithic farmers, and shorter than those of Caucasus and Western Hunter-Gatherers, suggesting that the inhabitants of Ganj Dareh did not undergo the large population bottleneck suffered by their northern neighbours. While some degree of cultural diffusion between Anatolia, Western Iran and other neighbouring regions is possible, the genetic dissimilarity between early Anatolian farmers and the inhabitants of Ganj Dareh supports a model in which Neolithic societies in these areas were distinct." - from "The genetics of an early Neolithic pastoralist from the Zagros, Iran" by Gallego-Llorente, et al, 2016
| Domesticating Plants
Cereal crops were first domesticated around 9,000 BC in the Fertile Crescent in the Middle East. The first domesticated crops were generally annuals with large seeds or fruits. These included peas, wheat and rye. The two most important grains domesticated were,
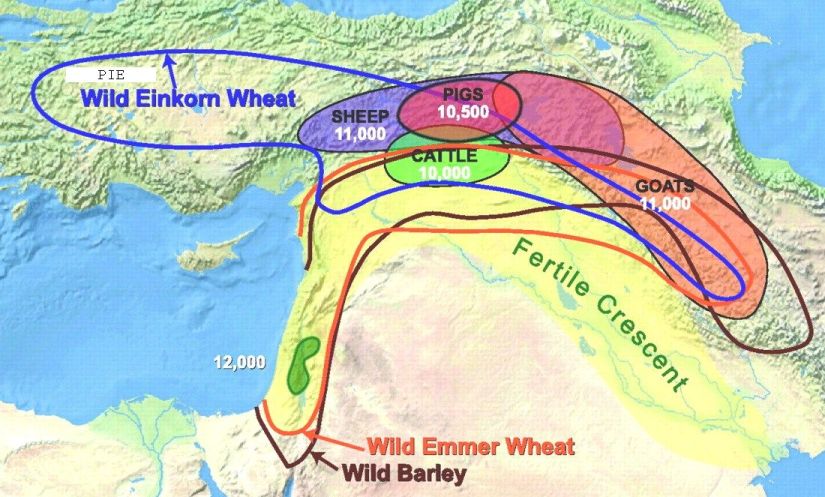 |
Domesticatiing Animals
The Dog Dogs were the first domesticated animals. This occurred 15,000-35,000 years ago, during the Paleolithic, and it is now thought that these early canines may have domesticated themselves. That is, while humans may have intentionally domesticated some wolves into dogs, for the most part wolves self-domesticated by establishing a mutually beneficial relationship with Paleolithic humans. They began by scavenging on the remains of prey animals left at human settlements or at kill sites. Those wolves that were less anxious around and aggressive to humans thrived and effectively colonized the human-dominated environment. Once this relationship was established humans began to use the canines for their own purposes. The domestication of other animals had to wait upon the Neolithic, when man established settlements that could hold the animals. Sheep, Goats and PigsBecause of their smaller size, these were the first food-source animals to be domesticated. Due to its aggressive personality, however, pigs trailed the others. Both Paleolithic and Neolithic man preyed on these animals, but it was only the latter that had the settled communities that made it possible to capture and hold some of these animals for later consumption. Feeding these captures and eventually breeding them followed.  Cattle
Cattle
Because of their size, and the danger this posed, cattle were the latest of the prey animals to be domesticated, which followed a similar course to the above. The Cat Cats first began to live in conjunction with humans during the Neolithic, drawn to farms in pursuit of the rodents that ate man's grain. They were, however, free-living, requiring nothing from humans and suffered to roam at will because of the service they provided. This reminds me of my father, a farm boy, who talked of the cats around the family farm. These cats were not treated as pets and certainly never fed, which would have defeated the purpose of them living there. They were not feral, but they also weren't tamed. The word is synanthropic, "a member of a species of wild animal or plant that lives near, and benefits from, an association with human beings and the somewhat artificial habitats that people create around themselves (see anthropophilia). Such habitats include houses, gardens, farms, roadsides and rubbish dumps." Cats followed human farmers as they moved into and across Europe. The Horse
|
The Neolithic Farming Revolution, continued

The invention of farming did not spring from a single source. Multiple cultures made this discovery at different times and in different places. Who was first is still open for debate. Recent research, ". . . reveals three genetically distinct farming populations living in the Near East at the dawn of agriculture 12,000 to 8,000 years ago: two newly described groups in Iran [Ganj Dareh] and the Levant [Natufians] and a previously reported group in Anatolia [Konya plain], in what is now Turkey. Together, the results suggested that agriculture spread in the Near East at least in part because existing groups invented or adopted farming technologies, rather than through population replacement . . . The findings tell a different story from what researchers believe happened later in Europe, when the first farmers moved in from Anatolia and largely replaced the hunter-gatherer populations who had been living there . . . The integration of ancient DNA samples from archaeological skeletal samples allows us to redefine the parameters of population history at the origin of agriculture. Clearly the transition from hunter-gatherer communities to agricultural ones is not a singular invention and spread, but the result of a series of innovations, aggregations and disseminations on both the cultural and biological sides of human history . . . The researchers also learned how descendants of each early farming group, even as they began to intermingle, contributed to the genetic ancestry of people in different parts of the world: Farmers related to the Anatolian group spread west into Europe, people related to the Levant group moved south into East Africa, people related to those in Iran or the Caucasus went north into the Russian steppe, and people related to both the farmers in Iran and hunter-gatherers from the steppe spread into South Asia." - from Anthrogenica.com Apparently there are only a handful of places around the globe that have native foods that could support such an innovation. The map below displays these places and the crop that supported them. Note that not all crops are equally valuable. While the cereals of the Middle East were easy to store and its lentils and peas were high in protein, the yams and bananas of New Guinea were much less nutritious and spoiled quickly, limiting the potential of the societies that depended upon them. Other factors that limited modernization were the availablity of suitable animals for domestication and navigable rivers to support trade. See "Guns, Germs, and Steel" by Jared Diamond for an excellent account of this. 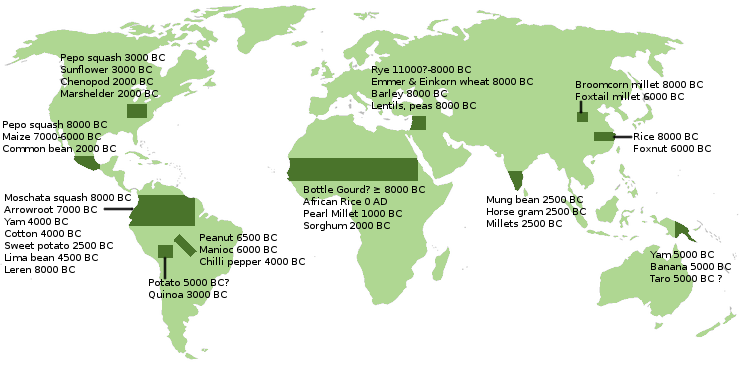 By the way, those environmentalists who believe that our capitalistic economy is unsustainable and whose battle-cry is "Back to the Paleolithic!," see the farming revolution as the great tragedy of human development. On a less extreme level, many dieticians look to the pre-Neolithic to identify foodstuffs more "natural" for the human diet than the carbohydrate-rich foods of the Neolithic farmers. That is perhaps true, but it takes a modern economy to make a Paleolithic diet possible for a large audience. |
Evidence that Anatolian hunter-gatherers developed farming on their own some 10,500 years ago has been found at Boncuklu hoyuk, on the Konya plain in central Anatolia. Stone tools, burned seeds, wheat chaff, and weeds known to have grown in early farmers' fields were discovered as were bones that suggested they kept sheep and goats. However, the tools at Boncuklu and other sites in central Anatolia are unlike those found at other early farming sites in the Fertile Crescent, suggesting that the Anatolians did not learn farming from the Levantines, but invented it on their own. That is, if farming had come to them through cultural diffusion out of the Levant, they would have used farm tools similiar to those of their teachers.
"In addition, the ancient DNA evidence now clearly shows that there is a distinctive local gene pool in the early Neolithic at places like Boncuklu, different from the genetics of Levantine Neolithic populations" - from Douglas Baird of the University of LiverpoolAgain, that is to say that it wasn't emigrants out of the Levant who physically moved into Anatolia, bringing farming technology with them. Anatolians who independently invented farming survived to pass their genes on to later Neolithic populations in central and western Anatolia. - from the Archaeologial Institute of America. Early farmers of the pre-pottery Neolithic had low genetic diversity.
As an update to the above, a new paper comments on the origins of the farmers of central Anatolia, to the west of the Anatolian farmers of the northern Fertile Crescent,
"Anatolia was home to some of the earliest farming communities. It has been long debated whether a migration of farming groups introduced agriculture to central Anatolia. Here, we report the first genome-wide data from a 15,000-year-old Anatolian hunter-gatherer [Pinarbasi] and from seven Anatolian [Boncuklu] and Levantine early farmers. We find high genetic continuity (~80-90%) between the hunter-gatherers and early farmers of Anatolia and detect two distinct incoming ancestries: an early Iranian/Caucasus related one [Caucasus Hunter Gatherer?] and a later one linked to the ancient Levant [Jordan and Israel] . . . Our results suggest a limited role of human migration in the emergence of agriculture in central Anatolia.So they are claiming they see a cultural diffusion of farming from the Fertile Crescent to Central Anatolia, unlike the late demic transfer seen in Europe. How do we know that the "two distinct incoming ancestries" didn't bring farming technology with them? It does seen clear, however, that the Anatolian Hunter-Gatherers picked up farming quickly, no matter the method of transfer, unlike their European peers.
. . . in regions of the Fertile Crescent such as the southern Levant and the Zagros Mountains (located between present-day eastern Iraq and western Iran), the population structure persists throughout the Neolithic transition, indicating that the hunter-gatherers of these regions locally transitioned to a food-producing subsistence strategy.
Central Anatolia has some of the earliest evidence of agricultural societies outside the Fertile Crescent and thus is a key region in understanding the early spread of farming. While archeological evidence points to cultural continuity in central Anatolia, due to the lack of genetic data from pre-farming individuals, it remains an open question whether and to what scale the development of the Anatolian Neolithic involved immigrants from earlier farming centers admixing with the local hunter-gatherers.
. . . We reveal that Neolithic Anatolian populations derive a large fraction of their ancestry from the Epipaleolithic Anatolian population, suggesting that farming was adopted locally by the hunter-gatherers of central Anatolia. We also detect distinct genetic interactions between the populations of central Anatolia and earlier farming centers to the east . . ." - from "Late Pleistocene Human Genome Suggests a Local Origin for the First Farmers of Central Anatolia" by Feldman, et al, 2019
Boncuklu hoyuk 8,300-7,500 BC
This very early Neolithic site is just 10 kilometers north of Catalhoyuk, see below. The people of Boncuklu lived in oval shaped buildings, usually dug down into the ground, with mud brick walls. This is quite unlike the pueblo-like buildings built at Catalhoyuk a thousand years later. Boncuklu Y-DNA samples are quite similar to the later farmers at Barcin, Mentese, and early European farmers. 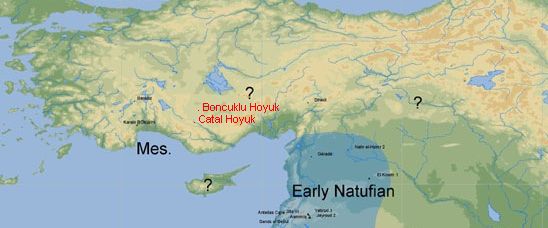 Asikli hoyuk 8,200-7,400 BC This Neolithic village is located north of Catal hoyuk on the bank of the Melendiz brook. Due to its date and structural organization it is known to be "a prime example of a first foray into sedentism." The village was located near an obsidian source, and became a base of trade which stretched to modern day Cyprus and Iraq. Wikipedia notes that some 6,000 obsidian objects were retrieved from the surface layer of the site. |
Why might our G2a forebears of Anatolia have begun their experiments in farming high, on the northern reaches of the Euphrates and Tigris rivers rather than lower, in what we might assume was the gentler course of these rivers? It is most likely because of the nature of these rivers and the plant and animal life that could be sustained in their various phases as they flowed to the sea.
It was only with the later invention of irrigation that the desert-like southern alluvial plains could be farmed."The rivers usually are discussed in three parts: their upper, middle, and lower courses. The upper courses are restricted to the valleys and gorges of eastern Anatolia, through which the rivers descend from their sources, lying 6,000 to 10,000 feet (1,800 to 3,000 metres) above sea level."
"Their middle courses gradually approach each other, bounding a triangle of mainly barren limestone desert known as Al-Jazirah (Arabic: 'The Island') in eastern Turkey, northern Iraq, and extreme northeastern Syria. There the rivers have cut deep and permanent beds in the rock, so that their courses have undergone only minor changes since prehistoric times. Along the northeastern edge of Al-Jazirah, the Tigris drains the rain-fed heart of ancient Assyria, while along the southwestern limit the Euphrates crosses true desert."
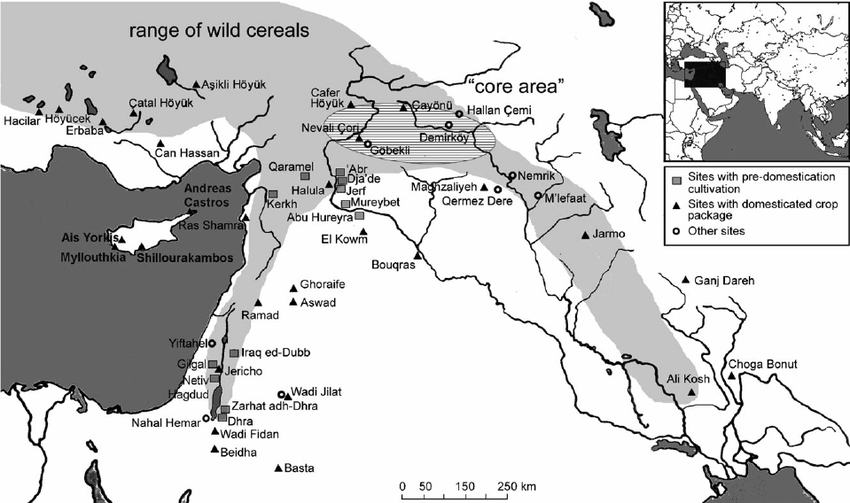
Ancient Y-DNA has been recovered at the following early Neolithic sites:
- Boncuklu hoyuk 8,300-7,500 BC, on the Konya plain of south central Anatolia, yielded G2a2b2b and G2a2b2b1a (6,500-6,200 BC).
- Tepecik-Ciftlik 7,500-5,800 BC, east of Boncuklu in the foothills of the Taurus mountains, yielded G2a2a (6,635-6,475 BC), as well as C1a2 and a probable J2.
- Barcin hoyuk circa 6,400 BC, near the Bosphurus, yielded G2a2a, G2a2b2a (P303), G2a2a1b, G2a2a1b1, G2a2b2a1c, G2a2b2a3, and G2a2a1a2a (6,500-6,200 BC), and G2a2b2a1a1c (6,067-5,982 BC), which is a sister clade of G-L497, as well as C1a2, H2, I2c and J2a.
- Mentese yielded G2a and G2a2a1 (6,400-5,600 BC), as well as I.
- Kumtepe circa 5,700 BC, southwesst of Barcin, G2a____
- Catalhoyuk, just south of Boncuklu, yielded G2a2b2b1a.
| Catalhoyuk
Or Catal Huyuk. A Neolithic town that, from 7,500 to 5,700 BC, was one of the world's largest settlements. At a time when most of the world's people were wandering hunter-gatherers, it had as many as 10,000 inhabitants living in its mud-brick houses. The pueblo-like settlement of Catalhoyuk, on the Konya plain in south-central Anatolia, was founded in the Neolithic by cereal and pulse [dry pea, lentil, chickpea, lupin and fava bean] farmers who also brought domesticated goats and sheep. Only a few centuries later (circa 6,500 BC) cattle were introduced to Catalhoyuk and other sites in Central Anatolia, presumably by trading with their eastern neighbours. Also around 8,500 years ago, G2a Neolithic farmers arrived in northwest Anatolia and Thessaly in central Greece, as attested by the ancient genomes sequenced by Mathieson et al. (2015) and Hofmanova et al. (2015). - derived from Haplogroup G2a 
|
| Gobekli Tepe
A ritual complex composed of as many as 20 circular temples containing curiously carved pillars, like that to the left, have been found. All the pillars are T-shaped with heights changing from 3 to 6 meters. Archeologists interpret those T-shapes as stylized human beings, mainly because of the depiction of human extremities that appear on some of the pillars. What also appears on these mystical rock statues are carvings of animals as well as abstract symbols, sometimes picturing a combination of scenes. Foxes, snakes, wild boars, cranes, wild ducks are most common. Most of these were carved into the flat surfaces of these pillars. There are also three-dimensional sculptures in the shape of a lion descending on the side of the T-pillar. The circular temples were probably covered by wood and thach roofs. A city subsequently grew up around the ritual complex and the needs of the growing population forced a more intensive form of food production. 
Alternate explanations of this site's settlement exist, of course. One is referred to as 'Beer before Grain.' It upends the idea that agriculture came first, followed by the discovery of brewing. The new theory says that people assembled at Gobekli Tepe to drink low-alcohol beer made from native wheat and barley. This quasi-religious bacchanalia was such a delight that peoplle rushed to find new sources of grain to increase the supply of beer, and the invention of farming followed. Another (less likely) theory is that Gobekli Tepe shows that mankind has gone through multiple rises & falls. The theory posits that this community was actually based on a farming technology (there is no proof for this) and they then built their great complex as a result of their new-found wealth, but crashed due to unknown factors, returning to a hunter-gatherer lifestyle. Another theory says aliens built the site (what would we do without aliens?), while another says its all a fraud perpetrated by the Turkish government. |
Another site of interest is Asikli hoyuk, in central Anatolia, just east of Catalhoyuk on the Konya plain. The major importance of the site is the role it played in the history of the region as early hunters and gatherers shifted from nomadic lifestyles to more permanent villages. A recent article indicates that both farming and animal domestication occurred here very early, ~8,350-7,500 BC. They determined this by the examination of salt deposits, indicative of heavy animal or human urine. Sheep and goats were amongst the earliest animals to be domesticated because of their small, managable sizes. Asikli Hoyuk was located near an obsidian source, and became a base of trade which stretched to modern day Cyprus and Iraq.
Based on the remains that have been found so far, the population of Anatolia during the Neolithic was 60% G2a and 30% H2. The following is an abstract that provides further background.
"It has hitherto been difficult to obtain genome-wide data from the Near East. By targeting the inner ear region of the petrous bone for extraction and using a genome-wide capture technology we achieved unprecedented success in obtaining genome-wide data on more than 1.2 million single nucleotide polymorphism targets from 34 Neolithic individuals from Northwestern Anatolia (~6,300 years BC), including 18 at greater than 1x coverage. Our analysis reveals a homogeneous population that is genetically a plausible source for the first farmers of Europe in the sense of (i) having a high frequency of Y-chromosome haplogroup G2a [as you'll read below, most Neolithic remains in Europe have been G2a], and (ii) low Fst distances from early farmers of Germany (0.004 +/- 0.0004) and Spain (0.014 +/- 0.0009). Model-free principal components and model-based admixture analyses confirm a strong genetic relationship between Anatolian and European farmers. We model early European farmers as mixtures of Neolithic Anatolians and Mesolithic European hunter-gatherers, revealing very limited admixture with indigenous hunter-gatherers during the initial spread of Neolithic farmers into Europe. Our results therefore provide an overwhelming support to the migration of Near Eastern/Anatolian farmers into southeast and Central Europe around 7,000-6,500 BC. Our results also show differences between early Anatolians and all present-day populations from the Near East, Anatolia, and Caucasus, showing that the early Anatolian farmers, just as their European relatives, were later demographically replaced to a substantial degree." - from "The Genetic Structure of the World's First Farmers" by Lazaridis, et al, 2016The last statement is significant and illustrates the limitation of trying to use modern DNA to explain ancient movements. The site exploited was Kumtepe, in the Troas of northwestern Turkey. This G2a population showed a strong continuity with that of present-day Sardinia. Mesolithic refers to a transition period between the Paleolithic and Neolithic.
| Kumtepe
Around 4,800 BC the first settlement in Kumtepe was founded. The inhabitants lived on fishing, and their diet included oysters. The dead were buried, but without grave gifts. Although Kumtepe belongs to Neolithic, the occupants used also copper. Around 4,500 BC the settlement was abandoned. Around 3,700 BC new settlers came to Kumtepe. The people of this new culture, Kumtepe B, built relatively large houses with multiple rooms, sometimes a porch. They also practiced animal husbandry and agriculture. The main domestic animals were goats and sheep, bred not only for meat but for milk and wool as well. They knew lead and bronze along with copper. Shortly after 3,000 BC Yassitepe and Hisarlik (Troy) were colonized probably from Kumtepe." - from Wikipedia |
Remains of the Tyrolean iceman, Otzi, and of those found in Chalcolithic [Copper Age] Remedello, in northern Italy, share a strong affinity with those from Kumtepe, as do those from Tepecik-Ciftlik, in south central Anatolia.
The Importance of Beer to Civilization
"Brewing and beer consumption have played a significant role in prehistoric human cultures around the world. Beer was a tasty, nutritious food, a substance affecting the mind, medicine, a religious symbol, as well as a social medium and an accelerator. Alcohol relieved the pain and prevented the spread of infection. Beer was a safe and healthy drink compared to contaminated water. At the time when our ancestors began to domesticate agricultural crops, they commonly produced not only bread but also beer. It is probable that the first ceramic vessels in the Near East were created precisely out of the need to more effectively control the technological process of beer production. Similarly, in the Central European Copper Age, beer production and its growing social significance influenced the emergence of the set Ceramic complex that lasted continuously for more than three millennia. Beer has entered almost all aspects of social life, from everyday consumption and social interactions to initiation ceremonies and major religious celebrations. The study of beer and other fermented beverages sheds light on many aspects of the biocultural development of humans on this planet." - from "Beer, Pottery, Society and Early European Identity" by Turek, et al, Jan 2020. The invention of bread and beer has been argued to be responsible for humanity's ability to develop technology and build civilization. People have been imbibing alcohol for most of their history in the form of rotting fruit, which ferments as it decays. Afterall, we know that apes, elephants and birds become drunk in this way. As hunter-gatherer tribes settled into agrarian civilizations based around staple crops like wheat and barley, they appear to have stumbled upon the knowledge that low-alcohol beer could be created from wheat and barley. This probably occurred when wild yeasts in the air caused spontaneous fermentation in grain products. In fact, some anthropologists have argued that these early peoples' insatiable thirst for hooch may have contributed to the Neolithic Revolution by inspiring new agricultural technologies. Little is known of this early invention because clay pots, which provide the best evidence we have of beer making, was not invented until about 2,000 years after the invention of agriculture. An alternative theory is that fermentation was discovered with native grains during the hunter-gatherer period, which led to the invention of farming to increase the available product for the increased manufacture of beer. |
No ancient G-type Y-DNA has yet been found amongst European remains that date to the Paleolithic. During this period it appears that 36% of its male population were part of haplogroup I and 64% were part of other haplogroups that have since gone extinct in Europe.
The Neanderthals, homo neanderthalis, had been living in Ice Age Europe for over 250,000 years when Cro-Magnon man, now more often known as early modern, or anatomically modern man, first started to advance into Europe from the Middle East. This migration began some 45,000 years ago and reached the Atlantic shore of western Europe about 10,000 years later. Competition between the two groups resulted in the extinction of the Neanderthal in Europe by about 24,000 years ago.
These early modern men were hunter-gatherers and mainly of haplogroups BT, CT, C1, F and IJK. Haplogroup I emerged later, about 17,000 years ago.
The number of these Paleolithic hunter-gatherers in Europe would not have been great; it takes a lot of territory to support a population that must live off the land. While it seems a surprisingly low number to me, it is estimated that no more than 70,000 humans lived in Europe at the end of the Paleolithic.
"From the Aurignacian to the Glacial Maximum, the metapopulation remained in a positive quasi-stationary state, with about 4400-5900 inhabitants (95% confidence interval (CI95%): 1700-37,700 inhabitants) . . . The metapopulation reached 28,800 inhabitants (CI95%: 11,300-72,600) during the mid-Late Glacial recolonisation." - from "Estimates of Upper Palaeolithic meta-population size in Europe from archaeological data" by Bocquet-Appel, et alA later study, however, estimated a population between 130,000 and 410,000 for this period - from "Human Population Dynamics in Europe over the Last Glacial Maximum" by Tallavaara, et al. Interestingly, this study was edited by Bocquet-Appel, the author of the citation above, so he must have accepted these new figures. However, that's still a remarkably low number for all of Europe.
A later study looked at the beginning of the Paleolithic, the Aurignacian period, and arrived at just 1,500 humans in Western and Central Europe,
"Demographic estimates are presented for the Aurignacian techno-complex (~42,000 to 33,000 y calBP) and discussed in the context of socio-spatial organization of hunter-gatherer populations. Results of the analytical approach applied estimate a mean of 1,500 persons (upper limit: 3,300; lower limit: 800) for western and central Europe." - from "Population dynamics and socio-spatial organization of the Aurignacian: Scalable quantitative demographic data for western and central Europe" by Schmidt, ZimmermannOf course, the population did increase over time, especially during the Mesolithic when the climate was friendlier and the people had better weapons. But these low population figures may explain why there is so little evidence of early Hunter-Gatherer ancestry in European Y-DNA. Geography is important too. Note that the Paleolithic Natufian society in the Levant had a relatively large population, but theirs was a paradise for hunter-gatherers. Europe was never able to support such large numbers until farming was introduced.
| Periods of Paleolithic Europe
See also Early Europe. Aurignacian43,000 to 33,000 years before the present (BP). Also known as Cro-Magnon, these Early Modern Humans (EMH) come out of the Middle East to spread across Europe. These have been described as a "robust" people, which means, I think, heavily built. a "The Aurignacian tool industry is characterized by worked bone or antler points with grooves cut in the bottom. Their flint tools include fine blades and bladelets struck from prepared cores rather than using crude flakes. The people of this culture also produced some of the earliest known cave art, such as the animal engravings at Trois Freres and the paintings at Chauvet cave in southern France. They also made pendants, bracelets, and ivory beads, as well as three-dimensional figurines. Perforated rods, thought to be spear throwers or shaft wrenches, also are found at their sites." - from WikipediaThe total human population in Europe was, at this time, extremely limited. Gravettian 33,000 to 22,000 years BP, although some elements lasted until 17,000 BP. Its disappearance roughly coincided with the bitter cold of the Last Glacial Maximum. As the climate chilled and glaciers advanced, this population had sought refuge in southern Europe, in Iberia and the Balkans. "The Gravettians were hunter-gatherers who lived in a bitterly cold period of European prehistory, and Gravettian lifestyle was shaped by the climate . . . Gravettian culture thrived on their ability to hunt animals. They utilized a variety of tools and hunting strategies. Compared to theorized hunting techniques of Neanderthals and earlier human groups, Gravettian hunting culture appears much more mobile and complex. They lived in caves or semi-subterranean or rounded dwellings which were typically arranged in small "villages". Gravettians are thought to have been innovative in the development of tools such as blunted-back knives, tanged arrowheads and boomerangs. Other innovations include the use of woven nets and oil lamps made of stone. Blades and bladelets were used to make decorations and bone tools from animal remains." - from WikipediaThese people were more "gracile" than the Aurignacian. Solutrean 22,000 to 17,000 years BP. The Solutrean industry is a relatively advanced flint tool-making style of the Upper Palaeolithic of the Final Gravettian and employed techniques not seen before and not rediscovered for millennia. Magdalenian 17,000 to 12,000 years BP. Later culture of the Upper Paleolithic and Mesolithic. "It was characterized by a cold and dry climate, the existence of humans in association with the reindeer, and the extinction of the mammoth. The use of bone and ivory as implements, begun in the preceding Solutrean epoch, increased, making the period essentially a bone period [i.e. not flint?]. Bone instruments are quite varied: spear-points, harpoon-heads, borers, hooks and needles . . . The later phases of Magdalenian culture are contemporaneous with the human re-settlement of north-western Europe after the Last Glacial Maximum." - from Wikipedia |
It was during the Neolithic period that our ancestors, haplogroup G2a, entered Europe from Anatolia, moving up through Greece and the Balkans into central Europe. They found a countryside that was very lightly settled.
What did these farmers from Anatolia look like? Examination of ancient remains indicates that they looked most like southern Europeans of today; Sardinian or perhaps Greek. They had dark hair and eyes, and a fairly dark complexion. The blonde haired, light skinned people of northern Europe did not develop until much later. Interestingly, the hunter-gatherers they met in Europe also had dark skin, but many had blue eyes.
"Skin pigmentation is a classic example of a polygenic trait that has experienced directional selection in humans. Genome-wide association studies have identified well over a hundred pigmentation-associated loci, and genomic scans in present-day and ancient populations have identified selective sweeps for a small number of light pigmentation-associated alleles in Europeans. It is unclear whether selection has operated on all the genetic variation associated with skin pigmentation as opposed to just a small number of large-effect variants . . . a large-effect variant at SLC24A5 was introduced to Europe by migrations of Neolithic farming populations but continued to be under selection post-admixture. This study shows that the response to selection for light skin pigmentation in West Eurasia was driven by a relatively small proportion of the variants that are associated with present-day phenotypic variation." - from "The evolution of skin pigmentation associated variation in West Eurasia" by Ju, Mathieson, et al"SLC24A5 appears to have played a key role in the evolution of light skin in humans of European ancestry." - from Wikipedia. SLC45A2 is also an important allele for determining skin tone.
 For many years researchers had debated how farming technology entered Europe. Was it via the transmission of the idea from one group to another, that is via cultural diffusion, or was it due to a mass migration of people, made dominant by their novel technology, displacing and replacing the existing hunter-gatherer groups they met along the way, the so-called demic diffusion? While cultural diffusion had long been the favored theory, the analysis of ancient DNA appears to have settled this question in favor of the latter.
For many years researchers had debated how farming technology entered Europe. Was it via the transmission of the idea from one group to another, that is via cultural diffusion, or was it due to a mass migration of people, made dominant by their novel technology, displacing and replacing the existing hunter-gatherer groups they met along the way, the so-called demic diffusion? While cultural diffusion had long been the favored theory, the analysis of ancient DNA appears to have settled this question in favor of the latter.
The record indicates that in about 7,000 BC Early Farmers (EF) of Anatolia began moving into Europe, taking with them their new technology. They displaced some of the Mesolithic peoples already living there, but apparently more often lived alongside them. In these early years the DNA record indicates that there was very little mixing between the native hunter-gatherers and the new farmers. The latter probably means that these new farmers brought their wives and families with them. But at least initially there was little motivation for social intercourse. The two cultures, one of wandering hunters and the other of sedentary farmers, had little in common to bring them together.
There was, however, conflict between the two societies. Most of the Neolithic sites so far discovered in Europe were fortified and exhumations of ancient remains often show signs of trauma most likely related to warfare. Hunter-gatherers, hungry during the long cold winters, would quickly learn that each farm village held stores of food that could soon be theirs if they just knocked a few heads. It is also true that because farming presupposed a sedentary lifestyle and an investment in improving the land, the control and defense of that property became important. What the hunter may have seen as fruit, for instance, free for the taking, was, to the farmer, a violation of his ownership rights. Their disputes were baked into their different view of land, not unlike later interactions between Europeans and American Indians.
While accepting and embracing progress seems obvious, here in the form of agriculture, farming is hard and hunting is easy, and often fun. I suspect few of the hunter-gatherers of Europe saw much advantage in settling down. The downside was a future risk, vague and uncertain. It took many years, and many risks to come to fruition, before the hunter-gatherers of Europe joined the farmers. Every winter or so a few hungry hunters would come to the gates of the fortified villages of the farmers to request entrance and acceptance. Another man to work the fields and man the walls would be welcome.
Over time the new European farmers picked up members from the local hunter-gatherers and integrated them into their communities. Examination of Neolithic remains discovered in Western Europe show a large number of haplogroup C1a and, especially, I2a had been added by the time the G2a farmers had migrated that far.
The earliest European Neolithic sites were in northern Greece and Thessaly. These sites were tested and contained G2a remains.
"Recent radiocarbon dating indicates that by 6,600-6,500 calibrated (cal) BC sedentary farming communities were established in northwestern Anatolia at sites such as Barcin, Mentese, and Aktopraklik C and in coastal western Anatolia at sites such as Cukurici and Ulucak, but did not expand north or west of the Aegean for another several hundred years. All these sites show material culture affinities with the central and southwestern Anatolian Neolithic. Early Greek Neolithic sites, such as the Franchthi Cave in the Peloponnese, Knossos in Crete, and Mauropigi, Paliambela, and Revenia in northern Greece date to a similar period." - from "Early Farmers from Across Europe directly Descended from Neolithic Aegeans" by Zuzana Hoftmanova, et alNote that at this early point the Black Sea may not yet have broken through the Bosporus, allowing a 'dry-shod' route to Europe. In addition there are thousands of islands in the sea between Turkey and Greece. While not all of them are inhabitable, you can, apparently, travel the Aegean without going outside the sight of some landmass. This would greatly facilitate the movement of peoples onboard the unreliable boats of this early era.
| Greek Neolithic
Early Neolithic sites in Greece are shown on the map to the right. The earliest villages are found in the eastern half of the country, from Macedonia in the north to the Peloponnese Peninsula in the south, with the largest concentration of villages in Thessaly, in central Greece. The area was not densely inhabited. There are probably no more than about three hundred Early Neolithic sites, and their total population at the time was no more than a few tens of thousands. The villages are found well inland, often near a copious spring or a perennial river. The early farmers apparently selected only the best and most reliable soils for farming. Studies of pollen from cores taken from lakes and swamps show that forest cover was not adversely affected by clearing of fields for nearly a thousand years after the first farmers arrived. Village farming was simple, based on the clearing of small fields that could be cultivated with digging sticks and hoes to grow wheat, barley, and pulses. Herds of cattle and sheep were grazed on meadows in nearby hills. The late Neolithic site at Kleitos, in Thessaly, yielded G2a2a1b (4,200-4,000 BC). "Genetic origins of the Minoans and Mycenaeans" by Lazaridis, et al, claimed that the ancient remains of 19 Bronze Age Minoan and Mycenaean individuals show that three-quarters of their ancestry came from the first Neolithic farmers of western Anatolia and the Aegean. |
Once established in northern Greece the Neolithic farmers began to follow two differenct routes further into Europe.
[The Northern Route] "G2a farmers from the Thessalian Neolithic quickly expanded across the Balkans and the Danubian basin, reaching Serbia, Hungary and Romania by 5,800 BC, Germany by 5,500 BC, and Belgium and northern France by 5,200 BC. Ancient skeletons from the Starcevo-Koros-Cris culture (6,000-4,500 BC) in Hungary and Croatia, and the Linear Pottery [LBK] culture (5,500-4,500 BC) in Hungary and Germany, all confirmed that G2a [included G2a2b with both cultures] remained the principal paternal lineage even after farmers intermingled with indigenous populations as they advanced."Those farmers who migrated west on the Southern Route moved more quickly than those that moved north through Central Europe. This was probably because, in moving directly west, they stayed in the same climate zone. Those farmers moving north transited zones of increasingly shorter summers and colder winters. They had to pause from time to time as they learned to cope with the new conditions.
[The Southern Route] "By 7,800 years ago, farmers making cardial pottery arrived at the Marmara coast in northwest Anatolia with ovicaprids [sheeps, goats] and pigs. These people crossed the Aegean by boat and colonized the Italian peninsula, the Illyrian coast, southern France and Iberia, where they established the Cardium Pottery culture (5,000-1,500 BC). Once again, ancient DNA yielded a majority of G2a samples in the Cardium Pottery culture, with G2a frequencies above 80% (against 50% in Central and Southeast Europe)." - from Haplogroup G2a
Note that pottery was developed and evolved with farming, providing the containers to safely store food stuffs over the winter. The nomadic hunter-gatherers had little need of such containers and less ability to carry them along. As the storage of excess food allowed people more free time, they began to decorate the pots in unique ways and passed these styles down to their descendants. It may be hard to imagine in our era in which a plethora of styles, an impossible variety of forms are available in such things as fabric and wallpaper. However, in this early time the style of your mother and grandmother was passed on directly to the next generation with little or no change attempted, or perhaps even allowed. Look at the wall decorations of Eqyptian tombs. Over a thousand years, in their cultural isolation, the style and the elements included were unvaried.
Ancient haplogroup G-type was discovered in Neolithic remains in the following Balkans locations:
- Dzhulyunitsa, Bulgaria, yielded G2a2b2b1a (6,300-6,150 BC) as well as C.
- Yabalkovo, Bulgaria, yielded G2a2a1a2a (6,000-5,900 BC) as well as I2a2.
- Govrievo, Northern Macedonia, yielded G2a2b2b1 (5,979-5,735 BC).
- Malak Preslavets, on the Danube river in northern Bulgaria, yielded G2a2b2a (P303) (5,800-5,400 BC) as well as T1a1 and C.
- Gomolava, Serbia, yielded G2a2a1 (5,604-5,376 BC) and a later set of G2a2a1 (4,710-4,460 BC).
- Cotatcu, Romania, yielded G2a2b2b (5,715-5,626).
- Ivanovo, Bulgaria, yielded G2a2b2a1a1c1a (4,725-4,605 BC).
- Varna, Bulgaria, yielded G2 (4,685-4,499 BC) as well as CT and R1.
| Old Europe
Old Europe is a term describing a relatively homogeneous pre-Indo-European Neolithic culture centered in the Danube river valley of southeastern Europe, but stretching as far as Britain. It occupied a time period from about 7,000 BC, when the first farming societies in Greece were founded, to 3,000 BC and the advent of the Bronze Age. Most European Neolithic groups shared certain basic characteristics. They lived in small-scale, family-based communities and were more egalitarian than the city-states and chiefdoms of the Bronze Age [assumed because they were small-scale?]. They subsisted on domesticated plants and animals supplemented with the collection of wild plant foods and hunting. They produced hand-made pottery without the aid of a potter's wheel. There were exceptions, of course, most notably the large cities of the Cucuteni-Trypillian. The survival of non-Indo-European languages, such as the modern Basque and the ancient Etruscan, are supposedly markers of this old culture. |
The farmers advance northward up the Balkans and into Central Europe was slowed as they passed through new climate zones.
"The Neolithic transition in Europe was driven by the rapid dispersal of Near Eastern farmers who, over a period of 3,500 years, brought food production to the furthest corners of the continent. However, this wave of expansion was far from homogeneous, and climatic factors may have driven a marked slowdown observed at higher latitudes. Here, we test this hypothesis by assembling a large database of archaeological dates of first arrival of farming to quantify the expansion dynamics. We identify four axes of expansion and observe a slowdown along three axes when crossing the same climatic threshold. This threshold reflects the quality of the growing season, suggesting that Near Eastern crops might have struggled under more challenging climatic conditions. This same threshold also predicts the mixing of farmers and hunter-gatherers as estimated from ancient DNA, suggesting that unreliable yields in these regions might have favoured the contact between the two groups." - from "Climate shaped how Neolithic farmers and European hunter-gatherers interacted after a major slowdown from 6,100 BCE to 4,500 BCE" by Betti, et al.These pauses in the Neolithic advance took place at specific times along each of the three tracks noted: 6,200 BC for Central Europe, 5,700 BC for northeastern Europe, and 5,400 BC for Scandinavia. All were, furthermore, linked to the fact that the climate in those regions was not, at those times, appropriate for the Middle Eastern crops the farmers brought with them. This had to do with the number of growing days of the proper temperature, not the severity of winter temperatures. Through a process of natural selection, hardier grain types evolved that eventually allowed the farmers to resume their advance into the north.
Interestingly, during these pauses in the advance there was an increase in admixture with WHG. Apparently during those periods of climatic pressure on food production, the farmers had need of the hunter gatherers' knowledge of where the best hunting and fishing were to be found, and so brought them into the fold. This latter may have helped in the increase in WHG haplogroups seen at this time.
| The Neolithic in Europe and the Spread of G2a
Haplogroup G has not been found in any Paleolithic or Mesolithic sites in Europe. Of Early/Middle Neolithic remains in Europe 61% are G2a, 16% I2a, 6% F, 3% C1a, 3% H2, 3% I, 3% T, 1.5% E1b1b, 1.5% I1, 1.5% J2, and 1.5% R1b - from eupedia.com. The Northern Route: 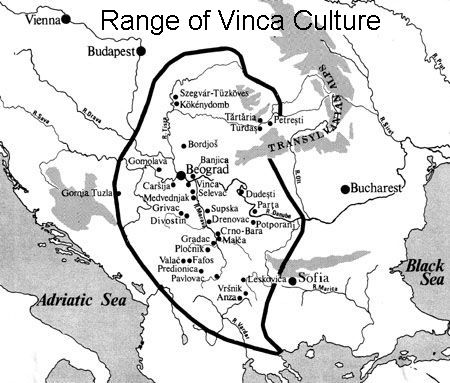 The Vinca Culture
The Vinca Culture
The earliest Neolithic culture to develop north of Greece was the Vinca. It ranged from northern Greece through Macedonia, Serbia, Montenegro, Bosnia, and parts of Romania and Bulgaria. "Farming technology first introduced to the region during the First Temperate Neolithic was developed further by the Vinca culture, fueling a population boom and producing some of the largest settlements in prehistoric Europe. The First Temperate Neolithic is an archaeological horizon consisting of the earliest archaeological cultures of Neolithic Southeastern Europe, dated to 6,400-5,100 BC. The cultures of the First Temperate Neolithic were the first to practice agriculture in temperate Europe, which required significant innovations in farming technology previously adapted to a mediterranean climate. - from Wikipedia.The Vinca practised a mixed subsistence economy with agriculture and animal husbandry combined with hunting and foraging. They introduced wheat, oat and flax to temperate Europe, and made increased use of barley. These innovations increased crop yields and allowed the manufacture of clothes made from plant textiles as well as animal products. There is indirect evidence that Vinca farmers made use of the cattle-driven plough. Two of three DNA samples taken from the ancient remains of the Vinca taken at Szederkeny-Kukorica-dulo, in southern Hungary, were tested as G2a2a and G2a2b2a1a (PF3346). The latter is part of the parent-stock of the Hissem family type. The other sample was H2-M282. Starcevo Culture Koros Culture Cris Culture
"Their Neolithic agricultural economy was based primarily on the cultivation of crops from the Fertile Crescent, such as Emmer wheat, Einkorn wheat, barley, spelt millet, pulses (peas and bitter vetch), and buckwheat. Some fruit trees were also cultivated, including plums and apricots. Starcevo farmers bred livestock, especially goats and sheep, but to a lower extent also cattle and pigs. They also supplemented their diets by fishing in rivers and hunting deer and wild boar in forests." - from EupediaGenetic testing of ancient Starcevo remains shows they were 57% G2a, 14% F, 14% I2a, 7% I, and 7% H2. See also, Starcevo-Koros-Cris Culture at eupedia.com. Cucuteni-Trypillian Culture
Four out of five Cucuteni-Trypillian samples from Ukraine were G2a2. The following seems to have been a mistake, based on a contaminated specimen [One of these, from Western Ukraine circa 3,619 to 2,936 BC, was G2a2b2a1a1b1a1a1, or G-L42, the earliest G-L497 found so far. That is, L42 is downstream from L497. Today there is still a hot spot of G-L497 in Moldavia. Note that L42 is a subclade of L43, which is a sister clade of our Z726. L42 is especially common today in Switzerland and the Austrian Tyrol, but it is also found in the Balkans and Italy. L42 seems to be associated with the Raetian-speaking region of Western Austria and Eastern Switzerland.] Linear Pottery Culture A successor to the Starcevo-Koros-Cris culture, located across Central Europe from Hungary to the Netherlands. Linear Pottery, more often known as LBK, from the German Linearbandkeramik, is a Neolithic decorative style that gets its name from the incised lines it bears, in imitation of painted bands. The pottery itself consists of simple cups, bowls, vases and jugs. "Farming was established in Central Europe by the Linearbandkeramik culture (LBK), a well investigated archaeological horizon, which emerged in the Carpathian Basin, in today's Hungary. However, the genetic background of the LBK genesis has not been revealed yet. Here we present 9 Y chromosomal and 84 mitochondrial DNA profiles from Mesolithic, Neolithic Starcevo and LBK sites (7th/6th millennium BC) from the Carpathian Basin and south-eastern Europe. We detect genetic continuity of both maternal and paternal elements during the initial spread of agriculture, and confirm the substantial genetic impact of early farming south-eastern European and Carpathian Basin cultures on Central European populations of the 6th-4th millennium BC. Our comprehensive Y chromosomal and mitochondrial DNA population genetic analyses demonstrate a clear affinity of the early farmers to the modern Near East and Caucasus, tracing the expansion from that region through south-eastern Europe and the Carpathian Basin into Central Europe. Our results also reveal contrasting patterns for male and female genetic diversity in the European Neolithic, suggesting patrilineal descent system and patrilocal residential rules among the early farmers. 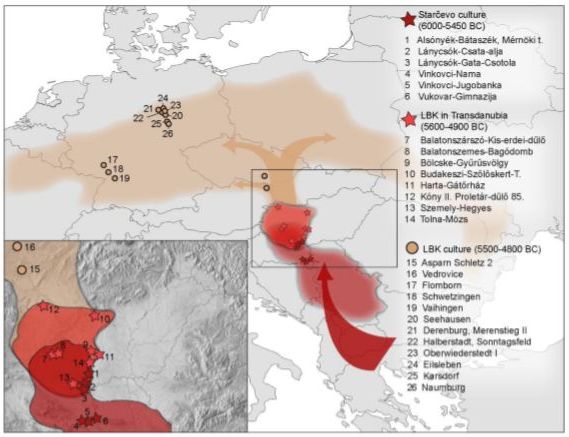
There is evidence of a violent relationship with the native hunter-gatherer population and, perhaps, other LBK communities as well. One-third of burials show evidence of traumatic injuries and a large number of LBK villages were fortified with ditches, walls and complex gates. "The later phase of the Central European Early Neolithic witnessed a rise in collective lethal violence to a level undocumented up to this date. This is evidenced by repeated massacres of settled communities of the Linearbandkeramik (ca. 5600-4900 cal BC), the first full farming culture in this area. Skeletal remains of several dozen victims of this prehistoric warfare are known from different sites in Germany and Austria. Here we show that the mass grave of Halberstadt, Germany, a new mass fatality site from the same period, reveals further and so far unknown facets of Early Neolithic collective lethal violence. A highly selected, almost exclusively adult male and non-local population sample was killed by targeted blows to the back of the head, indicating a practice of systematic execution under largely controlled conditions followed by careless disposal of the bodies. This discovery significantly increases current knowledge about warfare-related violent behaviour in Early Neolithic Central Europe." - from "Early Neolithic Executions Indicated by Clustered Cranial Trauma in the Mass Grave of Halberstadt" by Meyer, et alThe rise in collective violence to a level not seen before was probably because there was now a much larger, more concentrated population that possessed the resources to support greater levels of violence. So, life then was much as it is now, as violent as the resources allowed. The LBK culture flourished from about 5,500 to 4,500 BC. The greatest evidence for this group is along the middle Danube, upper and middle Elbe and the upper and middle Rhine. The burials at Derenburg Meerenstieg II in central Germany, tested as G2a2b2 (L141), are part of the LBK culture. They are dated to 5,500-5,000 BC. Genetic testing of ancient LBK remains shows they were 41.7% G2a, 16.7% F, 16.7% C1a2, 16.7% T1a, and 8% I1. Another Neolithic LBK site, at Karsdorf, Saxony-Anhalt, Germany (70 miles southeast of Derenburg) yielded 30 remains, dated 5,500-4,800 BC. This is south of Derenburg. 2 were T1, 22.2% were H2 from Derenburg and the remaining 55.6% were G2a from Halberstadt (just east of Derenburg) and Derenburg. All of these sites are in the north German plain. See also, Linear Pottery (LBK) Culture at eupedia.com. 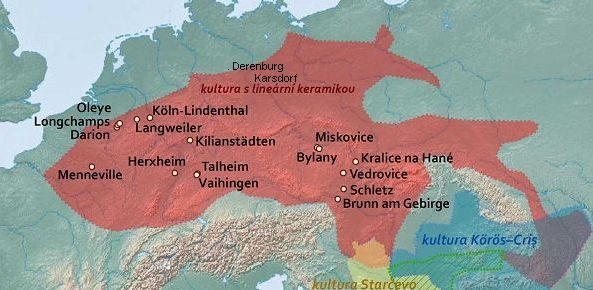 Funnel Beaker Culture
Salzmunde was the name of a late group, related to the Funnel Beaker culture, who resided in central Germany, 3,400-3,000 BC. "The analysis of two Funnelbeaker genomes from circa 3,000 BC using the Dodecad K12b calculator shows that they possessed over 60% of Atlantic_Med admxiture, more than any modern population except the Basques and the Sardinians, two populations with high Neolithic ancestry. This admixture is strongly reminiscent of the extent of the Atlantic Megalithic cultures." - from eupedia.comGenetic testing at Salzmunde shows they were haplogroups G2a2a and I2a1b1a. See also, Funnel-beaker Culture at eupedia.com. |
Ancient haplogroup G-type was discovered in Neolithic remains in the following Central European locations:
- Budakalasz, Hungary, yielded G2a2b2a1a (PF3346) (3,600-3,100 BC), and G2a2b2a1a1c1a (3,367-3,103 BC).Of some 102 Neolithic European remains, 41% were G2a, 41% I2a, 7% I/I1, 4% F, 3% CT/C1a, 2% H2, and 2% T.
- Schletz, Austria, yielded G2a2b2a3 (5,500-4,500 BC) - [G2a2b2a (P303) is an ancestor of our family type.]
- Halberstadt-Sonntagsfeld, north central Germany, yielded G2a2a (5,009-4,850 BC).
- Tyrol, Austria, yielded G2a2a1a2a1a (3,484-3,104 BC) - this was "Oetzi."
- Salzmuende-Schiebzig, central Germany, yielded G2a2a (3,400-3,025 BC) and I2a.
- Alsonnemedi, Hungary, yielded G2a2a (3,359-3,098 BC).
- Csajerszke, Hungary, yielded G2a2a1a2a1 (2,559-2,301 BC).
- Augsburg, Germany, yielded G2a2a1a2a1a (2,471-2,300 BC) and R1b.
- Szoreg, Sziv Utca, Hungary, yielded G2a2a1a2a (1,866-1,819 BC).
The Southern Route:
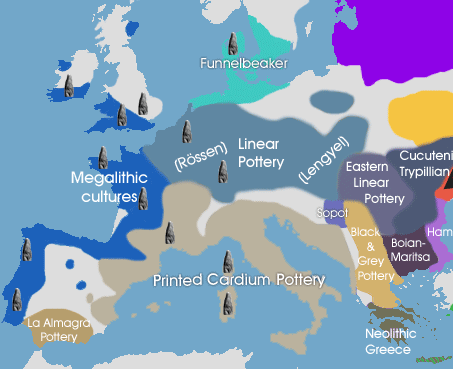
Printed Cardium Pottery Culture This pottery style gives its name to the main culture of the Mediterranean Neolithic. "Cardium Pottery or Cardial Ware is a Neolithic decorative style that gets its name from the imprinting of the clay with the shell of the cockle, an edible marine mollusk, formerly Cardium edulis, now Cerastoderma edule. These forms of pottery are in turn used to define the Neolithic culture which produced and spread them." - from WikipediaThis style of pottery is found from the Adriatic to the Atlantic coast of Portugal, and even into Morocco. The earliest ware, dating to about 8500 years ago, is found in western Greece, near Epirus and Corfu. The rapid spread of this culture suggests a seafaring expansion. The Avellaner cave burial site in Catalonia, Spain yielded G2a remains dated to about 5,000 BC. Genetic testing of other ancient Cardium culture remains shows they contained G2a, I2a1b1, E1b1b-V13, and R1b1c-V88. Lagozza-Chassey Culture The Late Neolithic Lagozza-Chassey, Chasseen-Lagozza, culture of France and northern Italy seems to have evolved from Cardium. These include the Treilles Group of southern France and the culture of Otzi, the frozen mummy of the Alps. Of 22 samples of ancient Y-DNA from Treilles 91% were G2a and 9% I2a. |
The testing of Neolithic remains in Europe seems to confirm, or at least strongly support, the idea that haplogroup G2a was the dominant lineage of Neolithic farmers and herders who migrated from Anatolia to Europe. It has been estimated that 60% of the European population was type G2a at this time. Substantial minorities of other haplogroups were found on different Neolithic sites next to G2a remains. These included haplogroups C, F and I, which may represent assimilated hunter-gatherers descended from Paleolithic Europeans. You could also surmise, I suppose, that these were the remains of slaves taken in conquest.
"From the start, however, G2a men were accompanied by a diversity of lineages assimilated from local hunter-gathering populations along the way, such as haplogroups C1a2, H2, E-M78, I*, I1, I2 and J2. Occasionally other Near Eastern lineages showed up, like one T1a sample in the LBK culture, and one R1b-V88 in northwest Spain. T1a tribes are thought to have domesticated ovicaprids [goats & sheep] in the Zagros mountains [border of Iran, Iraq and eastern Turkey], while R1b tribes would have domesticated cattle in the north of the Fertile Crescent." - from Haplogroup G2aA contrary paper was released in 2014, "Tracing the Genetic Origin of Europe's First Farmers Reveals Insights into Their Social Organization" by Anna Szecsenyi-Nagy, et al. It makes the following observations,
"Contrary to the low mtDNA diversity reported from hunter-gatherers of Central/North Europe, we identify substantially higher variability in early farming communities of the Carpathian Basin including the haplogroups N1a, T1, T2, J, K, H, HV, V, W, X, U2, U3, U4, and U5a. Previous studies have shown that haplogroups N1a, T2, J, K, HV, V, W and X are most characteristic for the Central European LBK [Linear Pottery Culture, see below] and have described these haplogroups as the mitochondrial 'Neolithic package' that had reached Central Europe in the 6th millennium BC. Interestingly, most of these haplogroups show comparable frequencies between the STA [Starveco], LBKT [Linear Pottery in Transdanubia] and LBK, comprising the majority of mtDNA variation in each culture."This paper claims that the Starcevo and LBK absorbed only a very few hunter-gatherer women and men. This may reflect a small Mesolithic hunter-gatherer population as discussed above.
"This is predominately based on a high number of 'Neolithic package' lineages and low frequencies of haplogroups attributed to hunter-gatherers, which clearly distinguish this cluster from hunter-gatherers of Central/North and southwest Europe, but also from Neolithic Iberian populations and Central European cultures of the 3rd/2nd millennium BC."
"Three STA individuals belong to the NRY [non-recombinant Y-chromosome] haplogroup F (M89) and two specimens can be assigned to the G2a2b (S126) haplogroup, and one each to G2a (P15) and I2a1."
"Considering the entire set of 32 published NRY records available for Neolithic Europe thus far, the low paternal diversity is indeed quite remarkable: G2a is the prevailing haplogroup in the Central European and Carpathian Basin Neolithic, and in French and Iberian Neolithic datasets . . . This very limited variation in NRY haplogroups in contrast to the high mtDNA haplogroups diversity suggests a larger effective population size for females than males. One plausible explanation for this phenomenon is patrilocality (where women move to their husband's birth place after the marriage). Other possibilities that could lead to similar observations include polygyny or male-biased adult mortality. A patrilocal residential rule was possibly linked to a system of descent along the father's line (patrilineality) in early farming communities. Ethnographic studies have suggested a change of residential rules at the advent of Neolithisation, showing different trends in residential rules among modern foragers and nonforagers. Increasing sedentism promotes territorial defence and control of resources, favouring men in the inheritance of land and property, which consequently led to patrilocal residence. At the same time, such residence pattern have to be momentarily flexible in expanding populations, allowing some of the sons to settle in new territories following population pressure and natural limitation of resources, e.g. after the carrying capacity of particular region has been reached. Patrilocality has also been raised in recent bioarchaeological studies. It has been suggested by aDNA evidence for the Treilles Neolithic community, and by stable isotope studies for the LBK in Central Europe"
 The early Neolithic sites located in Brunn am Begirge-Wolfholz were excavated between 1989 and 2005. The village, shown in an 18th century etching to the right, lies perhaps 20 kilometers southwest of Vienna. The site is comprised of about 75 longhouses in 5 separate groups and, for the entire settlement, date from 5540-5060 BC. In the oldest parts Linear Ceramic is missing, but the ceramics found were similar to Late Starveco. The later sites are clearly LBK. Current thinking is that the Neolithic settlers arrived from southern Hungary. Four remains from the site have been recovered and, of those, three yielded testable genetic material. All were male.
The early Neolithic sites located in Brunn am Begirge-Wolfholz were excavated between 1989 and 2005. The village, shown in an 18th century etching to the right, lies perhaps 20 kilometers southwest of Vienna. The site is comprised of about 75 longhouses in 5 separate groups and, for the entire settlement, date from 5540-5060 BC. In the oldest parts Linear Ceramic is missing, but the ceramics found were similar to Late Starveco. The later sites are clearly LBK. Current thinking is that the Neolithic settlers arrived from southern Hungary. Four remains from the site have been recovered and, of those, three yielded testable genetic material. All were male.
"Archaeogenetic research over the last decade has demonstrated that European Neolithic farmers (ENFs) were descended primarily from Anatolian Neolithic farmers (ANFs). ENFs, including early Neolithic central European Linearbandkeramik (LBK) farming communities, also harbored ancestry from European Mesolithic hunter gatherers (WHGs) to varying extents, reflecting admixture between ENFs and WHGs. However, the timing and other details of this process are still imperfectly understood. In this report, we provide a bioarchaeological analysis of three individuals interred at the Brunn 2 site of the Brunn am Gebirge-Wolfholz archeological complex, one of the oldest LBK sites in central Europe. Two of the individuals had a mixture of WHG-related [Western Hunter Gatherer] and ANF-related [Anatolian Neolithic Farmer] ancestry, one of them with approximately 50% of each, while the third individual had approximately all ANF-related ancestry. Stable carbon and nitrogen isotope ratios for all three individuals were within the range of variation reflecting diets of other Neolithic agrarian populations. Strontium isotope analysis revealed that the ~50% WHG-ANF individual was non-local to the Brunn 2 area. Overall, our data indicate interbreeding between incoming farmers, whose ancestors ultimately came from western Anatolia, and local HGs, starting within the first few generations of the arrival of the former in central Europe, as well as highlighting the integrative nature and composition of the early LBK communities."The G2a2 haplotype split into PF3146 and L30, the precursor of our G-L497. PF3146 then evolved into G2a2a1a-PF3177. Remains of the latter have also been found at Gomolava, Serbia (Vinca culture) and in the Maros region of southern Hungary (Koros culture).
. . .
"We obtained genetic data passing quality control for three out of the four individuals interred at Brunn 2. No usable genetic data was obtained from Individual 4. Individuals 1-3 were males by genetic typing. The mitochondrial lineages of Individuals 1-3 were J1, U5a1, and K1b1a (Table 1), while their Y chromosomal lineages were BT, CT, and G2a2a1a, respectively . . . SNP data obtained from whole genome sequencing were used as the basis for a PCA plot comparing to Neolithic Anatolian and early Neolithic European individuals (Figure 2). Individual 2 (I6913) fell closest to WHGs but shifted toward EEFs/ANFs, while Individuals 1 (I6912) and 3 (I6914) grouped with Anatolian Neolithic farmers and closely related central European farming groups. In the zoomed-in plot (Supplementary Fig. S1 online), I6914 appears to be borderline between ANFs and ENFs, while I6912 is on the high end of WHG relatedness for early European farmers. We note though that I6912 and especially I6913 have relatively low sequencing coverage, so their exact positions in PCA should be interpreted with caution."
. . .
"A bioarchaeological analysis of the remains of the interred at Brunn 2 presented in this study allows insight into the life history of early European Neolithic farmers who lived near the beginning of the establishment of farming economies in central Europe, revealing evidence of biological interaction between incoming Anatolians and local HGs during the earliest stages of the arrival of farming in the Neolithic Central Europe.
The mtDNA lineages of Individuals 1 and 3 belong to two of the most common mtDNA subclades found in Neolithic individuals from the Near East as well as their Neolithic European descendants. At the same time, an individual belonging to K1b (K1b2) has been identified in a Mesolithic forager from the Baltic. On the other hand, divisions of haplogroup U5 such as the U5a1 lineage identified in Individual 2 are generally considered to be characteristic of European hunter-gatherers. At the same time, a U5-carrying individual (U5b2) has recently been identified in the Catalhoyuk population of central Anatolia.
Individual 3 carries Y chromosome haplogroup G2a2a1a, from the larger set of G2a Y chromosomal lineages, which are characteristic of ANF and ENF populations. Individuals 1 and 2 have Y-chromosomes from the macro-lineages BT and CT, respectively, but due to their low-coverage data, we are not able to assign them with greater precision.
The genetic signature of Individual 3 is that of Neolithic Anatolian-related ancestry, consistent with that of most of the representatives of European Neolithic farming cultures, including LBK and Starcevo. Our analyses indicate that this individual had very little ancestry derived from European hunter-gatherers (likely zero, and no more than 1%). At the same time, Individuals 1 and 2 had WHG ancestry that was acquired after their ancestors had left Anatolia. We were unable to determine dates for this admixture, so it is possible either that it occurred locally or that the Brunn 2 migrants encountered WHGs along their journey and integrated WHG ancestry into the their predominantly ANF-derived genetic pool prior to their arrival in central Europe. It is also possible that the Brunn 2 migrants interacted with, or descended from, the Anatolia-derived farming communities (monochrome white painted pottery groups) that settled in the Balkans ca. 600 years earlier and would have also had opportunities to incorporate WHG ancestry in their gene pool since leaving Anatolia. However, the very high WHG-related proportion in Individual 2, combined with the western European affinity of the HG-related ancestry in Individual 1, points toward recent post-arrival admixture in central Europe as the most likely scenario."
. . .
"The absence of the evidence of violence at early LBK settlements suggests low hostility between the local HGs and LBK farmers."
- from "Interactions between earliest Linearbandkeramik farmers and central European hunter gatherers at the dawn of European Neolithization" by Nikitn, et al
It's interesting that the G2a individual, an ENF type, also had ENF mtDNA, K1b1a. The CT male, a WHG type, also had WHG mtDNA, U5a1, while the BT male, another WHG, had ENF mtDNA, J1. Quite a spread considering these were the only testable remains found.
Below are the frequency distribution, left, and variance distribution, right, for haplogroup G2a2b2a1a1b-L497. This is a European sub-clade and an ancestor of the Hissem/Heesom haplogroup. While the left-hand document fails to completely replicate the L497 frequency chart below [it was constructed by different people using a different data set I suspect], the right is interesting since it seems to show that the deep-source of the L497 sub-clade was in north central Germany. Remember, a large variance in subtypes indicates the region of the longest tenure of settlement. So, G-L497 may have first emerged in northern Germany and spread, both east and west and south. This area looks like the North German plain of Lower Saxony, a rich agricultural region. Ancient G2a remains, dated to 7,000 BP, were recovered nearby at the Derenburg Cave, a Linear Pottery cuture site. An article from 2013 states that G-L497 was created in central Europe and that 80% of the G type in the Alps is derived from this sub-clade - from "High resolution mapping of Y haplogroup G in Tyrol (Austria)" by Berger, et al, 2013.
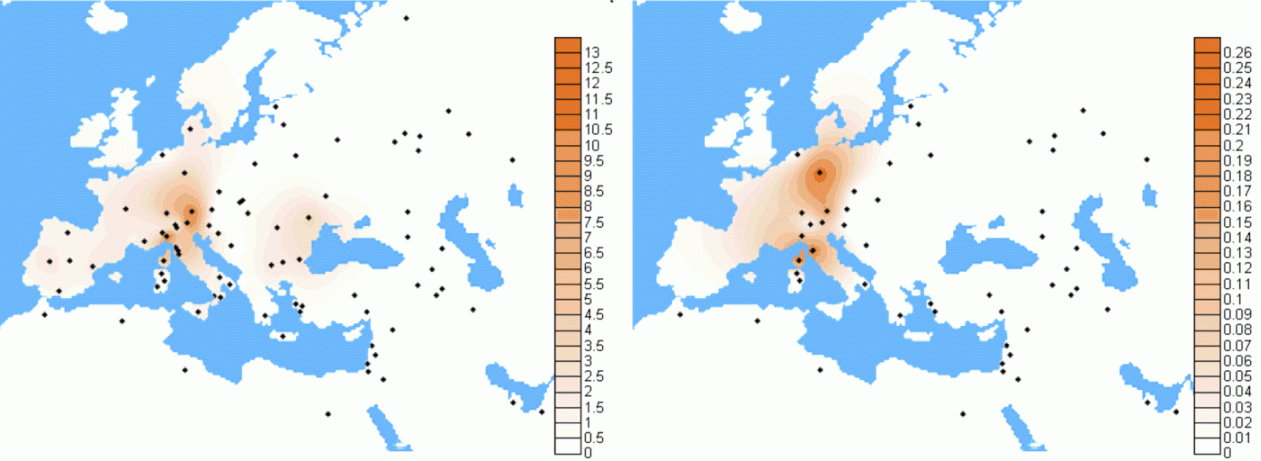
Our family Y-DNA type is G-Z726, which is downstream from G-L497. It may have arisen in southwest Germany.
| What Language Might these Neolithic People Have Spoken?
It is now generally accepted that the Indo-European language, on which almost all European languages are based, entered the continent after the Neolithic, during the Bronze Age. The one exception to this Indo-European rule is the Basque, who traditionally have lived in a region around the western end of the Pyrenees on the coast of the Bay of Biscay. Since they appear to be, autosomally, a blend of Mesolithic I2 and Neolithic G2a-dominant populations, perhaps their language would be a close approximation. The Etruscans are also thought to have spoken a non-Indo-European language, but the Empereor Claudius, circa 50 AD, is said to be the last person that could speak that language. |
Also of interest,
"G2a people may have been among the first humans to have acquired the alleles for fair skin. A hunter-gatherer from northern Spain tested by Olalde et al. 2014 still had dark skin as recently as 7,000 years ago. In contrast, an Early Neolithic farmer from Germany possessed the alleles for fair skin found in modern Europeans. The Neolithic individual was female, but Neolithic men from the same LBK culture were predominantly G2a. It is still unclear exactly when and among which haplogroup fair skin arose, but it has been suggested that the new diet brought by cereral agriculture would have caused deficiencies in vitamin D, which was traditionally absorbed from fish and meat among foragers. Mutations for light skin would have been positively selected among Neolithic agriculturalists to stimulate the production of vitamin D from sunlight in order to compensate for the scarcity of meat." - from Haplogroup G2a
So, to the best of my understanding, our direct line male-ancestors would have been busy farming a small plot some where in Central Europe, call it the German plain, at the end of the Neolithic, circa 3,000 BC . . . they would have had to make do with a stone plow point. It was not until some time later that they migrated to England. I base this on the estimation that the "father" of our family's terminal clade, G-Z726, evolved in Europe (modern examples are found across the continent) in about 2,600 BC, while its son, G-Z36217, is wholly a clade of English ancestry. It evolved circa 2,000 BC.

| The Chalcolithic or Copper Age
This was a transition period between the Neolithic and the Bronze Age. It began in about 5,000-4,500 BC when copper metallurgy was developed, either in the Middle East or the Balkans. This was a period before it was discovered that adding tin to copper created bronze, a far harder and stronger substance. Copper is one of the few metals that can occur in nature in a directly usable metallic form and may be found, in limited quantities, in pure form on the surface. Smelting refers to the extraction of metal from mined ore by a process of heating and melting. Casting refers to pouring molten metal into a mold and then allowing it to cool and solidify. It is thought that smelting and casting could have been invented in connection with high temperature ceramic kilns. By the late 5th millennium, there were four centers of metallurgy in the Near East: mountainous southeast Anatolia, an area in southeast Anatolia closer to the coast, the Levant west of the Jordan river, and highland Iran. It has been assumed that European metallurgy was derived from the Near East, however, based on carbon dating, it has been, "suggested that production of copper in the Balkans was an indigenous, independent, or nearly independent development...It would not be unreasonable to suggest that copper smelting began during the late sixth millennia BC [in the Balkans] and somewhat later in Anatolia. But, this conditional European priority depends on the resolution of slag like material from CCatalhoyuk level VIA, which would give priority back to Anatolia." - from "Copper Metallury-The Lost World of Old Europe-The Danube Valley-5,000-3,500 BC" by David AnthonyHowever, even if copper smelting was independently invented in the Balkans, and at an earlier date, there is no evidence that it spread from there to the Middle East. The next period of history recognized is the Bronze Age, staring in about 3,300 BC, when bronze metallurgy was developed. Bronze, a stronger metal, is made of copper alloyed with tin or arsenic. |
As we've discussed above, the family moved west into Europe over many centuries. The first chart below, from Scaled Innovation, shows longitude, that's the line on the globe stretching from north to south, versus time to visualize this migration. Note that moving up on this chart is equal to moving west. That the family was already as far west as the Alpine longitude by 8,000 BC seems a bit premature. I thought G2a was no further west than central Turkey at that time. The Scaled Innovations website does note that, "SNPs with lower testing levels are the most scattered: paths from T, G, and J2 tend to be poorly determined." Note also the area I've circled. That is the longitude for an area between the Rhine river to the east, and the Atlantic coast (I've noted Calais), to the west, where, presumably, the Hissem family lived from about 3,500 to 2,500 BC. That would be the time period when the Z726 SNP emerged. Now that looks just about right to me.
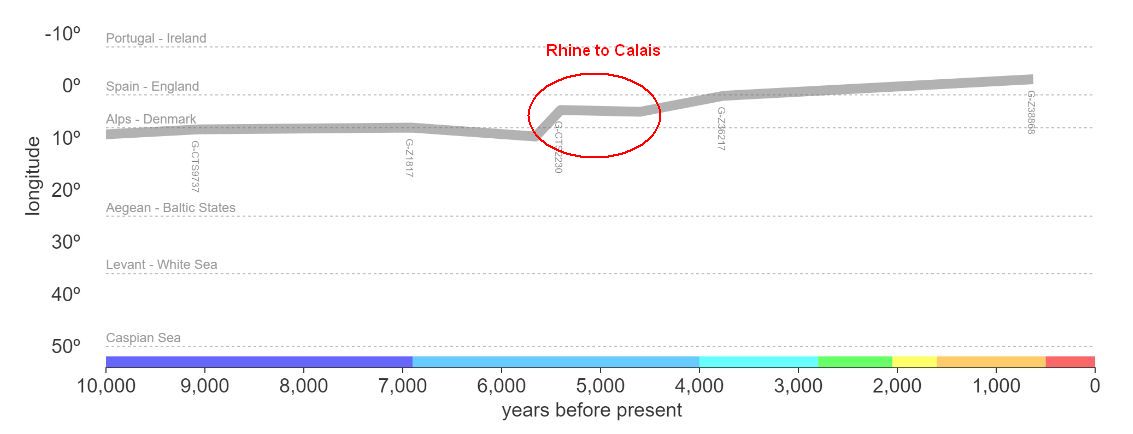
How fast did they move? The second chart, also from Scaled Innovation, shows how many kilometers per year they averaged. Note that the time-scale at the botttom is logrithmic to allow the whole period to be shown with enough clarity in the most recent past. The speed of the movement into Europe around 8,000 BC is notable. I guess that illustrates that for most of history people had little need or desire to move. Then, with the advent of agriculture, a fire got lit under people to find new lands for their crops to support their burgeoning populations.
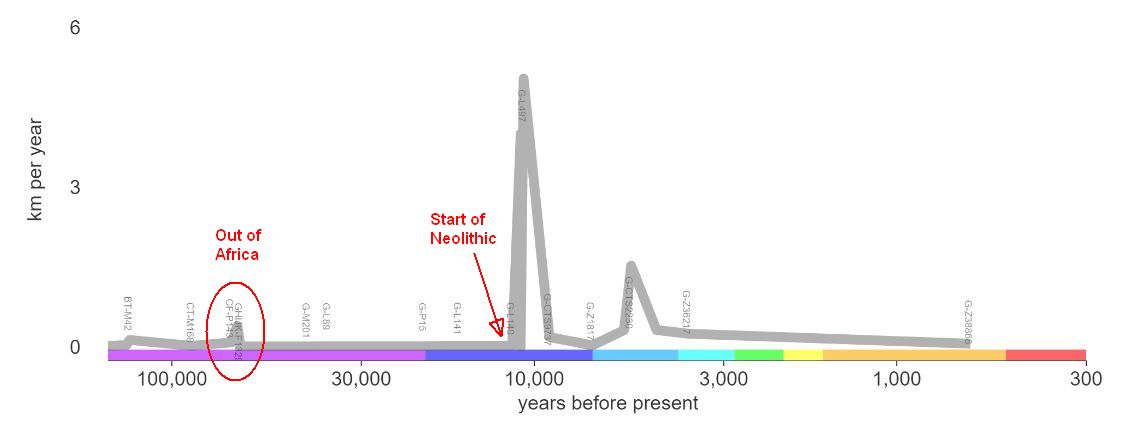
Now let's return to the locus of our search, England.
The Peopling of EnglandThe island of Britain has been settled by members of the homo genus on at least eight separate occasions, but they were driven back the first seven times by succeeding glacial periods that made the island uninhabitable. Based on excavations performed in and around Heysham, we know that the current occupation of Lancashire by modern man, homo sapiens, began only about 12,000 years ago, at the end of the last glaciation. Before that time a great glacier, a mile thick, extended as far south as Wales and the Midlands. The British people, then, are relatively late arrivals to their current homeland.
"Australian aboriginals have been in Australia longer, continuously, than the British people have been in Britain. There were probably people in the Americas before 12,000 years ago," - from Dr. Chris Stringer, London Natural History Museum
The last glaciation period began 2.5 million years ago. After the glaciers receded, but before the return of normal temperatures, the coastline was six to nine miles further west and a landbridge connected England to the continent.
| The Last Glacial Period
Within ice ages there are cycles of advancing and retreating glaciers caused by variations in global temperature. The last period of increasing cold and advancing glaciers within the current ice age, known in England as the Devensian glacial, lasted from about 110,000 to 12,000 years ago. The maximum extent of glaciation in this period occurred approximately 22,000 years ago. Sheets of ice 1,500 meters thick covered northern England and Europe, and sea levels fell dramatically because so much water was trapped in the ice. To the east, the Caucasus and the mountains of Turkey and Iran were capped by local ice sheets. The warmer periods within ice ages, when glaciers are retreating, are known as interglacials and, during the current ice age, these cycles have recurred at 40-100,000 year intervals. We are today at the peak temperature of the latest interglacial, the Holocene, which began about 12,000 years ago. The great issue of the global warming debate is, has man caused global temperatures to spike beyond what should have been expected at this point in the warming cycle. |
"The island of Britain suffered some of the most extreme climate changes of any area in the world during the Pleistocene." - from Dr. Chris Stringer, London Natural History Museum
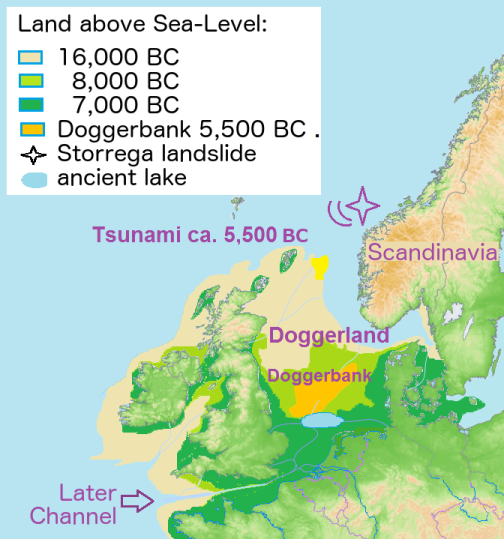 The Land Bridge
The Land Bridge
During the coldest period of the last glacial maximum large amounts of water were taken up by the glaciers. As a result the sea level fell. This revealed a large landmass connecting England with the continent. This has been called Doggerland, for the current-day Dogger Banks, a relatively shallow area in the North Sea. When temperatures began to rise the glaciers began their slow retreat, shedding water back into the oceans and causing the sea level to rise again. The area near the face of the glaciers continued to be inhospitable, having a tundra-like climate not unlike Arctic Russia or northern Alaska today. The people of Europe, the early homo sapiens, had, during the coldest period, retreated to sanctuaries in the south. They would have been slow to return, but, following the herd animals who were their prey, they were pulled north. Doggerland continued to exist for some time, only slowly being engulfed by the sea. As late as 7,000 BC a land bridge still allowed movement across by people and animals. Doggerland was finally swallowed and England's population was isolated, at least until ship construction became adequate to face the English Channel. The latest theory posits that a tsunami, caused by a tremendous landslide in Norway, called the Storegga Slide, destroyed the last vestiges of the land bridge in a single monumental wave. |
The first true men to arrive on the island were Paleolithic hunters coming across the land bridge, probably following the herds they preyed upon. As the climate continued to warm they moved further north. Early man's presence in Lancashire was proven by the discovery of the Poulton Elk. The remains of this animal, which was what we'd call a Moose in North America, were dated to about 10,000 BC. It was found at Poulton-le-Fylde, south of Heysham, on the Wyre river. Before dying it had been wounded 17 times by the stone weapons of Paleolithic hunters - with the primitive weapons available in that era it took a lot of effort to take down a large animal. Apparently the elk escaped into a wetland where the hunters could not follow, then perished there.
It must still have been mighty cold at this time in Lancashire.

These hunters were not our ancestors. So far, the remains found of this era have been haplogroup I2a.
It is not yet known when G2a people first got to England, though they may have been part of the Neolithic wave that reached the eastern shore of the English channel by about 4,000 BC. Note, however, that no haplogroup G2a remains have yet been found at Neolithic sites in England.
"So far there are NO Gs from Ancient Britain - even from the Neolithic. All of the Neolithic Britons were I2." - from Ted Kandell at the Facebook Haplogroup G pageThat does not mean G2a wasn't there, absence of evidence is not evidence of absence, but their presence has not been proven. The haplogroup that was in Britain in the Neolithic, and that exclusively, was I2a, which is so often found with G2a at continental sites.
The Neolithic arrived fairly late in England, over two thousand years after it first arrived in Europe. Where did this Neolithic influence arrive from? There are two obvious routes, the northern up the Danube river (G-P303) and the southern through the Mediterranean (G-M406).
- The Northern Route: The middle-Neolithic Rossen culture, which spread across northern France, the Low Countries, and central Germany, was a successor to the Linear Pottery culture which was itself 41.7% haplogroup G2a. However, while
"[i]t is suggested the late Rossen culture may be ancestral to the Neolithic cultures of Britain and Ireland (a group of cultures previously known as Windmill Hill culture), but there is no great similarity in the form of houses or pottery. According to alternative theories, the British Neolithic culture(s) came from Brittany." - from WikipediaThe Neolithic in Brittany came out of the south.
| Rossen Culture
"It has been suggested that there is a genetic continuity between LBK and Middle Neolithic populations including those associated with the Rossen culture." - from "Ancient DNA Insights from the Middle Neolithic in Germany" by Esther J. Lee, et alAlso, "They [the Rossen] provided a substantial background to the expansions of the Neolithic both to the British Isles and possibly also into northwest Europe." - from "Encyclopedia of Indo-European Culture" edited by J.P. Mallory and Douglas Q. AdamsThe Rossen and Lengyel were associated with haplogroups G2a, I2, I2b, and, to a lesser extent, J1 and T. |
"We assembled genome-wide data from six Mesolithic and 67 Neolithic individuals found in Britain, dating from 10.5-4.5 kBP, a dataset that includes 22 newly reported individuals and the first genomic data from British Mesolithic hunter-gatherers. Our analyses reveals persistent genetic affinities between Mesolithic British and Western European hunter-gatherers over a period spanning Britain's separation from continental Europe. We find overwhelming support for agriculture being introduced by incoming continental farmers, with small and geographically structured levels of additional hunter-gatherer introgression. We find genetic affinity between British and Iberian Neolithic populations indicating that British Neolithic people derived much of their ancestry from Anatolian farmers who originally followed the Mediterranean route of dispersal and likely entered Britain from northwestern mainland Europe." - from "Population Replacement in Early Neolithic Britain" - from Brace et alSee also,
"British Neolithic farmers were genetically similar to contemporary populations in continental Europe and in particular to Neolithic Iberians, suggesting that a portion of the farmer ancestry in Britain came from the Mediterranean rather than the Danubian route of farming expansion." - from "The Beaker Phenommenon" by Olalde et al, 2017So Mesolithic, or late Paleolithic Britains were similar to the hunter-gatherers of Western Europe. This is expected since the land bridge to Britain was still available in the early Paleolithic. The surprise is that the Neolithic was spurred by an influx of Iberian farmers who came west from Anatolia via the Mediterranean. That would mean they were chiefly G2a of the M406 sub-clade, undoubtedly accompanied by haplogroup I2a (whose Neolithic remains have been found in Britain). However, G-M406 is currently extremely rare in England. That may mean that these original farmers were replaced at some point and that the G-L497 population that exists in England today came into the island during the Bronze Age or later.
As has been noted above, ancient Y-DNA of the Atlantic Megalithic culture (5,000-2,000 BC) has been recovered for sites in Western Europe, though not in England. Tests yielded haplogroups CT (1 sample), G2a (20 samples), H2 (1 sample), and I2 (40 samples). It is not unreasonable to believe that some of these G2a peoples of this culture, as well as the I, would also reside across the channel in England. In a recent paper looking at burials at megalithic sites, all testable male samples were haplogroup I2a.
| Atlantic Megalithic Culture
The Megalithic culture began during the Neolithic, circa 5,000 BC, and flourished until about 2,000 BC, then going into a decline to end circa 1,500 BC. 
|
Neolithic farming in England was initially concentrated on the chalk and limestone plateaux of the Wessex Downs and the Cotswolds. These early farming communities were centred on large earth settlements, the remains of which can still be seen on many hill-tops in Dorset and Wiltshire. Here, the loamy, calcareous soils were sufficiently deep and fertile to support regular cropping, but were naturally well-drained and relatively easily cleared.
"Around the period between 3,500 and 3,300 BC, agricultural communities had begun centring themselves upon the most productive areas, where the soils were more fertile, namely around the Boyne, Orkney, eastern Scotland, Anglesey, the upper Thames, Wessex, Essex, Yorkshire and the river valleys of the Wash. These areas saw an intensification of agricultural production, and larger settlements." - Wikipedia
By 3,500 BC the Windmill Hill culture of the Salisbury Plain in southern England had developed. Its main occupations were the farming of wheat and barley, and raising cattle, sheep, goats, and pigs. Their culture shows similarities to the Cortaillod of the Swiss Lakes of western Switzerland, which was contemporaneous with the Rossen.
It took about 2000 years for farming to spread across the British Isles - from the BBC History website. The practice would have been difficult around Heysham due to the generally poor soil. However, a large number of polished axe heads from the Neolithic period were found there. Flint is not found in this district so the axes and hammers of the Stone Age were made of Bluestone. The remains of a Neolithic long house, built circa 3,500 BC, were found in Lancashire.
So far all of the Neolithic remains discovered in England have been tested as haplogroup I2. This group emerged in the Paleolithic. Present-day haplogroup G percentages in English counties are shown in the table below, extracted from "Genetic History of the Bristish and the Irish" at eupedia.com, dated October 2016. It shows percentages of the population having haplogroup G in the areas denoted. The fact that Lancashire and Cumbria both have such low numbers, while the northeastern counties and East Anglia have the highest tends to indicate an east-to-west migration vice the north-to-south I've used on the Roman Origins page. The latter assumed a Roman auxillary soldier as our ancestor, a Raetian, stationed at Hadrian's Wall, then drifting south after the collapse of the Empire.
Present-day Haplogroup G Percentages in English Counties- East Anglia includes data for Cambridgeshire, Norfolk and Suffolk
East Anglia: 4%
Northeast England: 3%
Wales: 2.5%
Cornwall and Devon: 2.5%
Southwest England: 2%
West Midland: 1.5%
Home Counties: 1.5%
East Midlands: 1%
Cheshire and Lancashire: .5%
Cumbria: 0%
In the Late Neolithic there was a resurgence of Western Hunter Gatherers (WHG) and a boom of haplogroup I2 to the detriment of Anatolian Farmers lineages like G2, T and H2. This appears to be associated with a climate change that stressed the agricultural economy.
"The Neolithic decline was a rapid collapse in populations between five and six thousand years ago . . . While heavily-populated settlements were regularly created, abandoned, and resettled during the Neolithic, after around 5400 years ago, a great number of those settlements were permanently abandoned. The population decline is associated with worsening agricultural conditions and a decrease in cereal production. Other suggested causes include the emergence of communicable diseases spread from animals living in close quarters with humans [i.e. the bubonic plague]." - from WikipediaThat the initial decline of the farmer population of Europe occurred due to possibly the earliest evidence of bubonic plague is per David Reich. See How One Small Tribe Conquered the World 70,000 Years Ago
The climate seems to have been so bad that in the British Isles and most of Scandinavia cereal production nearly ceased. Crop growing came back to the British Isles in the Bronze Age because new varieties of wheat had been developed which could be planted in spring and harvested in fall (spring wheat), unlike the original varieties which were planted in autumn and harvested in the spring (winter wheat). Those varieties couldn't grow in the cold, wet winters of the Isles.
| The Bronze Age
This era has classically been said to begin in about 3,300 BC and is characterized by the smelting of copper and tin, or arsenic, to create a harder metal, known as bronze. However, bronze artifacts have recently been found in the Balkans securely dated to 4,650 BC confirming the indepedent discovery of this technique in Europe. However, production lasted for only 500 years, ending with the collapse of the culture. Bronze was reintroduced to Europe by a migrating population circa 3,000 BC. Central Europe would not enter the Bronze Age until 1,800 to 1,600 BC while the Atlantic coast had to wait until 1,300 to 700 BC. The next period of history recognized is the Iron Age, starting in the 12th century BC. The last defined era is the Hisotrical when written records first became available in, perhaps, about 500 BC. |
In about 3,000 BC haplogroup G2a, the dominant Neolithic group in Europe, suddenly became relatively rare. This appears to have been due not to some widespread disease or cataclysm of nature, but to an invasion of Europe by riders out of the east who brought with them a new technology, bronze tools and weapons, and a new mode of transportation, the horse.
According to current hypotheses, it appears that migration into Europe occurred in three distinct waves,
- Paleolithic. After the Last Glacial Maximum, when glaciers began their slow retreat from Europe, a Paleolithic hunter-gatherer population of homo sapiens, comprised of haplogroups C1a/b, F, H and I, repopulated Europe, expanding out of southern refuges where they had fled during the worst of the cold.Now for an authority to say the same thing,
- Neolithic. Then, during the early Neolithic, about 6,000 BC, these people were joined by early farmers migrating out of Anatolia comprised predominantly of haplogroup G2a. They took two routes, one westward along the Mediterranean coast, the other through northern Greece, up the Hungarian plain, and into central Germany. They were the first in Europe to use pottery (hunter-gatherers didn't need it) and founded what has become known as the Cardium and Linear pottery cultures.
- Bronze Age. Later, about 3,000 BC, these groups were subsumed in a massive invasion by R1a and R1b populations coming out of Asia. This included the Yamnaya of the Russian steppe. Riding the first domesticated horses in Europe and wielding bronze weapons, they would have been formidable foes. Disease may have also played a role in their conquest.
"We described the individuals in our dataset in terms of their genetic relatedness to a hypothesized set of ancestral populations, which we refer to as their genetic ancestry. It has previously been shown that the great majority of European ancestry derives from three distinct sources: first, 'hunter-gatherer-related' ancestry that is more closely related to Mesolithic hunter-gatherers from Europe than to any other population, and that can be further subdivided into 'eastern' (EHG) and 'western' (WHG) hunter-gatherer-related ancestry; second, 'northwestern-Anatolian-Neolithic-related' ancestry related to the Neolithic farmers of northwest Anatolia and tightly linked to the appearance of agriculture; and third, 'steppe-related' ancestry that appears in western Europe during the Late Neolithic-to-Bronze Age transition, and which is ultimately derived from a population related to Yamnaya steppe pastoralists. Steppe-related ancestry itself can be modelled as a mixture of EHG-related ancestry and ancestry related to Upper Palaeolithic hunter-gatherers of the Caucasus (CHG) and the first farmers of northern Iran." - from "The Genomic History of Southeastern Europe" by Mathieson, et al, 2018
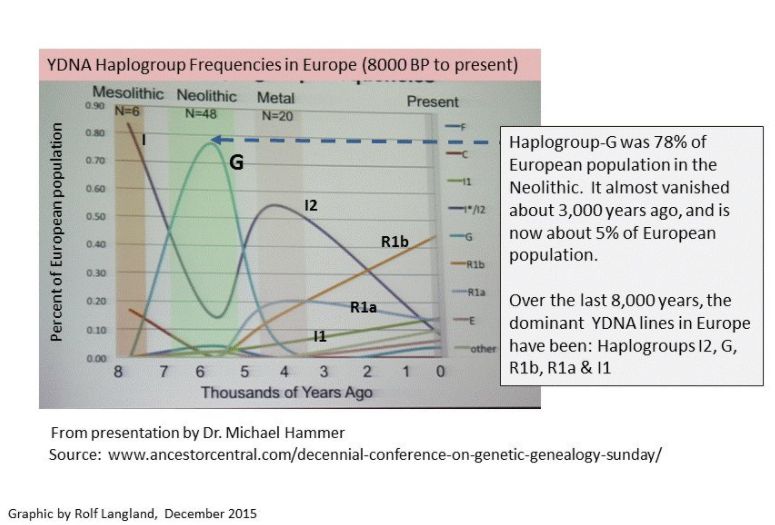
The disease theory for the collapse of haplogroup G2a is not dead, at least not as an adjunct to the invasion theory.
"A team of researchers led by scientists at the Max Planck Institute for the Science of Human History has sequenced the first six European genomes of the plague-causing bacterium Yersinia pestis dating from the Late Neolithic to the Bronze Age (4,800 to 3,700 years ago). Analysis of these samples, published in Current Biology, suggests that the Stone Age Plague entered Europe during the Neolithic with a large-scale migration of people from the Eurasian steppe."While this might explain the extremely rapid decline of male Paleolithic and Neolithic groups during this period, it does not explain the continued presence of ancient female lines.
In "Ancient Genomes Reveal Social and Genetic Structure of Late Neolithic Switzerland" by Furtwangler, et al, they had analyzed,
"96 ancient genomes from Switzerland, Southern Germany, and the Alsace region in France, covering the Middle/Late Neolithic to Early Bronze Age. Similar to previously described genetic changes in other parts of Europe from the early 3rd millennium BCE, we detect an arrival of ancestry related to Late Neolithic pastoralists from the Pontic-Caspian steppe in Switzerland as early as 2860-2460 calBCE. Our analyses suggest that this genetic turnover was a complex process lasting almost 1000 years and involved highly genetically structured populations in this region."
"In individuals older than 2600 BC originating from the sites of Oberbipp, Aesch and Muttenz only haplogroups belonging to the clades I2a and G2a were found."
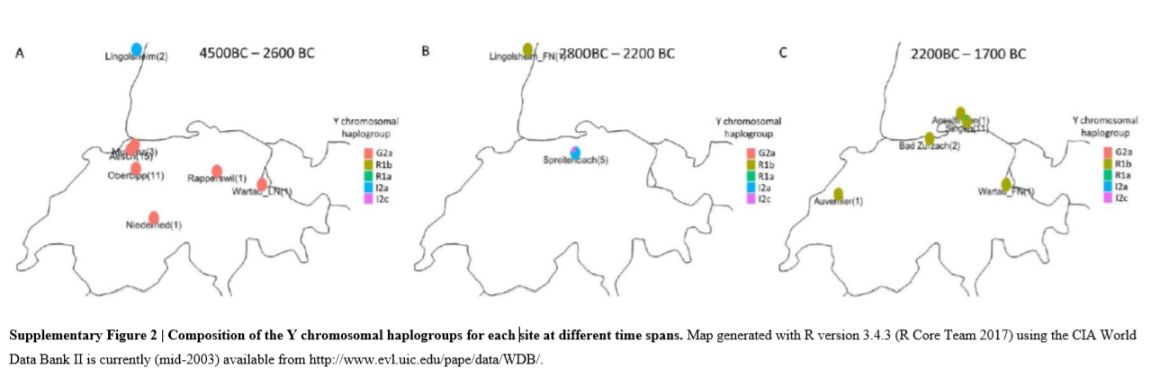
ID.........Site............Date BC....Term Mut..YHG*This may be an entry error because PF3239 is G2a2a1a2a1. G2a2b2a1a1 is PF3345, which represents the majority of all G in modern day Europe.
Aesch01 Aesch (CH) 3090-2917 PF3239 G2a2a1a2a1
Aesch12 Aesch (CH) 3010-2884 PF3239 G2a2a1a2a1
Aesch13 Aesch (CH) 3016-2901 PF3239 G2a2a1a2a1
Aesch14 Aesch (CH) 3014-2898 PF3239 G2a2a1a2a1
Aesch17 Aesch (CH) 3011-2889 PF3239 G2a2a1a2a1
Aesch19 Aesch (CH) no collagen PF3239 G2a2a1a2a1
Aesch20 Aesch (CH) 2913-2878 FGC7739/Z6488 G2a2a1a2a
Aesch21 Aesch (CH) no collagen PF3239 G2a2a1a2a1
Aesch22 Aesch (CH) 2892-2694 PF3239 G2a2a1a2a1
Aesch23 Aesch (CH) 2881-2676 PF3239 G2a2a1a2a1
Aesch24 Aesch (CH) 2912-2877 FGC7739/Z6488 G2a2a1a2a
Aesch4 Aesch (CH) 3094-2926 PF3239 G2a2a1a2a1
Aesch6 Aesch (CH) 2905-2759 PF3147 G2a2a
Aesch7 Aesch (CH) no collagen FGC7739/Z6488 G2a2a1a2a
MX150 Oberbipp Horgen (CH) 3244-3102 L91/PF3246/S285 G2a2a1a2
MX182 Oberbipp Horgen (CH) 3338-3031 PF3239 G2a2a1a2a1
MX183 Oberbipp Horgen (CH) 3344-3037 FGC7739/Z6488 G2a2a1a2a
MX187 Oberbipp Horgen (CH) 3337-2908 PF3239 G2a2a1a2a1
MX204 Oberbipp Horgen (CH) no collagen FGC7739/Z6488 G2a2a1a2a
MX209 Oberbipp Horgen (CH) no collagen PF3239 G2a2a1a2a1
MX210 Oberbipp Horgen (CH) no collagen PF3239 G2a2a1a2a1
MX211 Oberbipp Horgen (CH) 3100-2928 PF3239 G2a2a1a2a1
MX212 Oberbipp Horgen (CH) 3323-2581 PF3239 G2a2a1a2a1
MX213 Oberbipp Horgen (CH) 3363-2930 PF3239 G2a2a1a2a1
MX219 Oberbipp Horgen (CH) 3330-3216 PF3147 G2a2a
MX298 Wartau (CH) 2620-2448 P303 G2a2b2a, a precursor to L497
MX299 Oberbipp Horgen (CH) 2910-2679 PF3147 G2a2a
RA58 Muttenz (CH) 2937-2886 FGC7739/Z6488 G2a2a1a2a
RA61 Muttenz (CH) 2905-2865 PF3239 G2a2a1a2a1
RA62 Muttenz (CH) 2921-2886 PF3239 G2a2a1a2a1
SX10 Rapperswill Zürichstrasse (CH) new-born or unborn baby of SX8 L166 G2a2a1a2a1a
SX11 Niederried Ursisbalm (CH) 4458-4362 PF3239 G2a2b2a1a1*, the immediate precursor of L497
| Wartau
On the eastern edge of Switzerland. There are a number of Mesolithic, Neolithic and Bronze Age sites. Niederried UrsisbalmIn the Interlaken district of central Switzerland. The oldest trace of a settlement in the area is a Neolithic grave in Ursisbalm and a La Tene grave at Stadeli. |
Before we move on, it must be said that the horse itself may be the most significant reason for the success of the R1a/b invasion out of Asia. Not only did the horse provide an unrivaled speed of advance, but the very riding of such large, powerful beasts would have been unnerving to a victim population that had never seen such a thing. It might have seemed almost magical. Imagine your response to an army racing forward riding on the backs of analagous beasts, like bulls and bison [think of Mongo riding into town on a Brahma bull in "Blazing Saddles"]; overpowering, unbelievable, and panic inducing. An analog is the Aztecs' frenzied response to Spanish soldiers on horse-back. The Aztecs thought horse and rider were a single beast, like some mad centaur.
This large migration out of the Pontic-Caspian steppe may have been caused, as so many other things are, by climate change. Beginning in about 3,200 BC the climate in Europe grew colder and drier. About the same time the hitherto successful Cucuteni-Trypillian culture of the western Ukraine collapsed, perhaps due to drought induced crop failure. The pastoralists of the steppe moved west into this void, themselves possibly driven by the lack of good forage in those bad times.
While the R1a/R1b movement into Europe was characterized above as an invasion, it was a slow-motion one that took hundreds of years to reach the Atlantic coast. The horse-riders moved, settled, grew, and then moved on in stages across the continent.
Unlike the early farmers migrating out of Anatolia, these people quickly mixed with the native population, probably indicating that this was predominantly an invasion of single men who took wives from wherever they settled.
"Europeans who were alive from before the Yamnaya migration inherited equal amounts of DNA from Anatolian farmers on their X chromosome and their autosomes, the team reports today in the Proceedings of the National Academy of Sciences. This means roughly equal numbers of men and women took part in the migration of Anatolian farmers into Europe.Some have conjectured that this indicates that the original Yamnaya population was polygamous, and that it was the younger sons, lacking access to women of their own age, were forced to move west to obtain for both land and spouses. Could be . . .
But when the researchers looked at the DNA later Europeans inherited from the Yamnaya, they found that Bronze Age Europeans had far less Yamnaya DNA on their X than on their other chromosomes . . . the team calculated that there were perhaps 10 men for every woman in the migration of Yamnaya men to Europe (with a range of five to 14 migrating men for every woman). That ratio is extreme, even more lopsided than the mostly male wave of Spanish conquistadores who came by ship to the Americas in the late 1500s, Goldberg says." - from "Thousands of Horsemen may have swept into Bronze Age Europe, Transforming the Local Population" by Ann Gibbons in Science of 21 February 2017
Paleolithic haplogroups C, F and H disappeared from Europe. Haplogroup I was pushed into northern Germany and Scandanavia, and southeast into the Balkans.
The Neolithic G2a population retreated into highland sanctuaries. The number of G2a survivors plummeted in the subsequent years. Where they had once made up 50-60% of the Neolithic population, they now were only 5% of the total. If the R1a/b invaders took their wives from the local population that probably explains the dramatic drop in the next generation of Neolithic men. Looking at a map of the present day distribution of G-L497, the ancestor of the Heesom/Hissem sub-clade, which includes modern-day dispersions, there are interesting concentrations that remain in three areas: the Alps of Switzerland, Austria and northern Italy, with an associated population in Tuscany; today's Moldavia, remnants, I believe, of the Cucuteni-Trypilllian culture; and northeastern Iberia. These clusters are, I believe, traces of the refuges of our ancestors.
"Nowadays G2a is found mostly in mountainous regions of Europe, for example, in the Apennine mountains (15 to 25%) and Sardinia (12%) in Italy, Cantabria (10%) and Asturias (8%) in northern Spain, Austria (8%), Auvergne (8%) and Provence (7%) in south-east France, Switzerland (7.5%), the mountainous parts of Bohemia (5 to 10%), Romania (6.5%) and Greece (6.5%). The hilly terrain of southern Europe indeed makes it ideally suited for herding goats, which G2a men brought with them during the Early Neolithic period. But the most likely explanation is that mountains provided refuge for G2a tribes after the Proto-Indo-European speakers invaded Europe from the steppes of Russia and Ukraine during the Late Copper Age and the Bronze Age" - from eupedia.comOther researchers have been more outspoken, claiming something like genocide on the part of the R1a/R1b invaders. That seems extreme to me based on the very limited data we're working with.
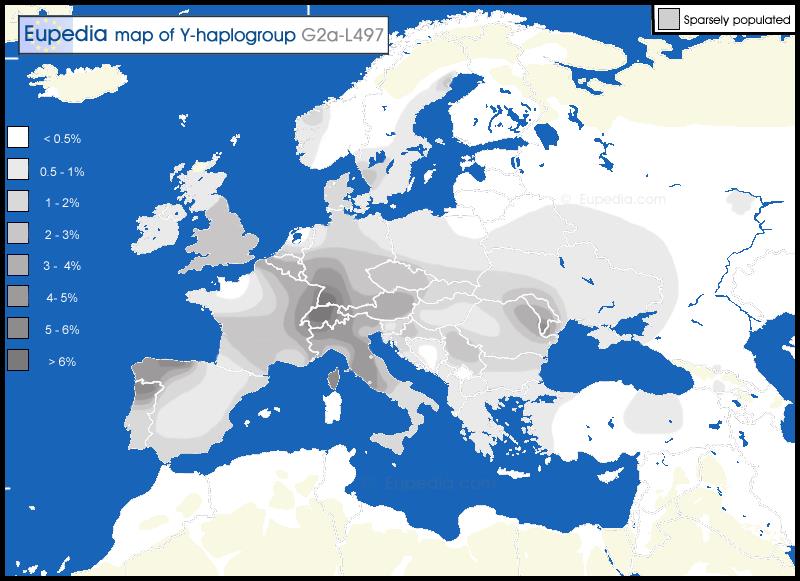
An interesting footnote to the story of the decline of haplogroup G2a in Europe is the fate of haplogroup I2a. While it was pushed to one side during the Bronze Age, it is today the most common paternal lineage in the former Yugoslavia, Romania, Bulgaria and Sardinia, and a major lineage in most Slavic countries. Why the different fate?
The following is a jump to my current theory of when the ancestors of the Hissem/Heesom family moved to the island of Britain. See The Bell Beaker Culture.
The following traces the evolution of haplogroup G based on the 2020 ISOGG haplogroup tree. The basal type emerged out of a transitional haplogroup, GHIJK, a daughter of haplogroup F.
Haplogroup G, a mutation at M201, emerged from haplogroup F in the Fertile Crescent of the Middle East in about 46,500 BC, per the Yfull Haplogroup G Tree [40,000 BC per Scaled Innovation]. At first, expansion of the type was slow, painfully slow, reflecting the very small initial population . . . well, one to begin with, but not truly flourishing for some time. Yfull employs a shorthand method for figuring TMRCA (Time to Most Recent Common Ancestor) utillizing the date of emergence of the daughter clades; the TMRCA for G-M20a would be 23,200 BC.
| TMRCA
A measure of how long ago any two male individuals likely shared a common patrilineal ancestor. The TMRCA for any two men can be determined by testing and comparing Y-STR marker profiles. The strength and accuracy of the TMRCA prediction increases as more Y-STR markers are tested and compared. |
Haplogroup G2, a mutation at L89 [P287], emerged in about 23,200 BC, per Yfull [35,000 BC per Scaled Innovation], also in the Middle East. This was almost 25,000 years after the basal type evolved. The designations shown as [ ] are where Yfull.com or the ISOGG differs from FTDNA in nomenclature.
Haplogroup G2a, a mutation at P15, emerged in about 18,600 BC, per Yfull [18,000 BC per Scaled Innovation]. The rate of mutation has slowly ramped up with just 5,000 years for this SNP to evolve.
Haplogroup G2a2, a mutation at L1259 [CTS4367], is the parent of G2a2a and G2a2b. It evolved in about 16,600 BC, per Yfull [15,000 BC per Scaled Innovation].
Haplogroup G2a2b arose as a mutation at L30 [CTS574]. It is the largest subgroup of G2a2. It evolved in about 14,500 BC, per Yfull [15,000 BC per Scaled Innovation]. Yfull, oddly, has CTS574 as a child of L30 while FTDNA and ISOGG list it as an equivalent variant. ISOGG also lists L30 as the parent of G2a2b1, a mutation at M406, while Yfull lists it as a sister clade of P303.
Haplogroup G2a2b2 arose as a mutation at L141 [CTS2488]. It evolved in about 12,400 BC, per Yfull [14,000 per Scaled Innovation], still in the Middle East. It came through Anatolia and up the Danube valley through the northern route described above. It is found in west and central Europe (see the remains found in Derenburg & Karlsdorf on the north German plain) and also amongst the Brahmins of India.
Haplogroup G2a2b2a, a mutation at P303. It evolved in about 12,400 BC, per Yfull [12,000 per Scaled Innovation], in the Middle East. It is heavily concentrated in Europe west of Russia and the Black Sea, but is still found in Turkey, Iran, and the Caucasus. I've recently seen discussions that P303 broke off from L141 at the Younger Dryas, 12,900 to 11,700 years ago, when farming first developed in the Pre-Pottery Neolithic A, see above. Note also the discussion above about the Anatolian Neolithic farmers of southwestern Turkey; P303 had a huge expansion during the first stage of this early period.
"The incredibly rapid population expansions in G2a2a-PF3147 and G2a2b2a-P303 happened between 12,700 and 10,800 years ago. In particular, the tMRCAs of the multiple *parallel* branches right under G-P303 (7 branches) and G-PF3177 (3 branches) starting at 11,500 years ago coincides with the modal earliest radiocarbon dates for Gobekli Tepe at 11,400 calBP (9,400 calBC)." - from Ted KandellGobekli Tepe was primarily a religious site associated with the Pre-Pottery Neolithic-A. It is located in southeastern Turkey. It does not seem to have been a settlement in the normal sense and no testable remains have been found at the site to date.
Ancient remains of G2a2b2a-P303 have been discovered at Barcin hoyuk, in northwestern Turkey, see the map to the right. These have been dated to 6,500-6,200 BC. Barcin hoyuk is north of Kumtepe, near the Sea of Marmara. Note that the landbridge from Anatolia to Europe still existed at this time. P303 defines the group that migrated into Central Europe.
G-M406, a sister clade of L141 [CTS2488], above, defines the group that migrated into Southern Europe. It is found in the Caucasus, but in more significant numbers in Turkey, Greece and in the eastern Mediterranean countries of Syria, Jordan and Lebanon, history's Phoenicia. It has not been found in Paleolithic DNA.
"In contrast to its widely dispersed sister clade defined by P303, hg G-M406 has a peak frequency in Cappadocia, Mediterranean Anatolia and Central Anatolia (6-7%) and it is not detected in most other regions with considerable P303 frequency. The expansion time of G-M406 in Anatolia is 12,800 years ago, which corresponds to climatic improvement at the beginning of the Holocene and the commencement of sedentary hunter-forager settlements at locations . . . thought to be critical for the domestication of crops (wheat and barley) that propelled the development of the Neolithic." - "Distinguishing the co-ancestries of haplogroup G Y-chromosomes in the populations of Europe and the Caucasus" by Siiri Rootsi, et alG-M406 arrived in Europe through the Mediterranean, along the southern route described above. An M406 population persists in Sicily, Sardinia and Corsica, though often in the more remote regions of those islands, where the inhabitants fled in the face of subsequent invasions by Phoenicians, Greeks, Etruscans, Carthaginians, Romans, Byzantines, Normans and Arabs.
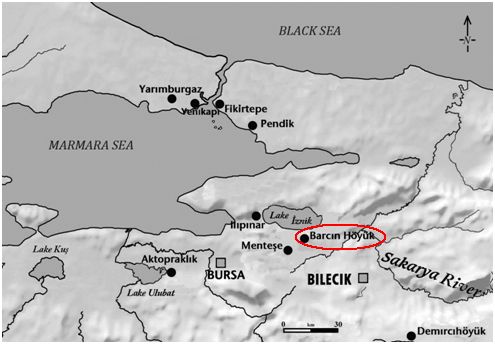
Haplogroup G2a2b2a1 is a mutation at L140. It evolved in about 9,500 BC, per Yfull [9,600 BC per Scaled Innovation], in the Middle East. This was an era marking the beginning of the farming revolution of the Neolithic. It is a geographically diverse group found in both western Asia, India and Europe, where almost 80% of the G-group are of this type. This type was found in remains found in Gurgy, on the Yonne river, in eastern France.
Early Neolithic farmers moved north and west from Anatolia through Thessalonika in northern Greece and into the Pannonian plain of Serbia and Hungary. The latter hosted the Starcevo-Koros-Cris culture of farmers circa 6,200-4,500 BC. "Their Neolithic agricultural economy was based primarily on the cultivation of crops from the Fertile Crescent, such as Emmer wheat, Einkorn wheat, barley, spelt millet, pulses (peas and bitter vetch), and buckwheat. Some fruit trees were also cultivated, including plums and apricots. Starcevo farmers bred livestock, especially goats and sheep, but to a lower extent also cattle and pigs. They also supplemented their diets by fishing in rivers and hunting deer and wild boar in forests." - from Eupedia.comNote that the Yamnaya migration into Europe, though it carried mainly R1b, also contained G-L140 that had been assimilated while they were still the steppe, just north of the Caucasus.
Haplogroup G2a2b2a1a is a mutation at PF3346. It evolved in about 8,800 BC, per Yfull [9,000 BC per Scaled Innovation], probably somewhere in Anatolia, though some push this into Europe. This type was found in ancient remains taken from Szederkeny-Kukorica-dulo, in southern Hungary, circa 5,230 to 5,050 BC. This was during the era of the Vinca culture, see above.
Haplogroup G2a2b2a1a1 is a mutation at PF3345. It evolved in about 8,600 BC, per Yfull [9,000 BC per Scaled Innovation], probably in western Anatolia or northern Greece.
"During the Neolithic and Chalcolithic, lots of subclades of haplogroup G proliferated across Europe. Now, only one of these subclades really thrives (G2a2b2a1a1 PF3345), representing the majority of all G in modern day Europe. What were the keys to its reproductive success?"
"DNA data suggests dates and places for early PF3345 that correspond with the Neolithic Cucuteni Tripolye culture of Romania and Moldova, with two successful branches developing during the 4th millennium BC - 1. An Eastward expansion (branches of U1) towards the Caucasus, 2. A more general Westward migration towards Southern Germany.
As these developments seem to mirror those of R1b-L23 (R1b-Z2103 in the East and R1b-U152 in the West), both in terms of dates and locations, perhaps PF3345 thrived by forming a minor component of some Yamnayan and Bell Beaker populations?
Bell Beaker mitochondrial DNA also has its strongest match with Cucuteni Tripolye samples, suggesting some degree of mixture between these R1b-L23 and G-PF3345 peoples.'
"I have identified four strands of G-CTS342 that moved into Italy; my estimates are that each of these strands originated to the North, mostly likely Bavaria. My date estimates for the moves of these four strands each fall between 1500 and 2500 BC, which ties up closely with similar estimates for the expansion of R1b-U152 from Southern Germany into Italy.
As R1b-U152 Bell Beaker also has its closest mtDNA match with G-PF3345 Cucuteni Tripolye, and as both U152 and CTS342 appear to have flourished at roughly the same time, I would predict that R1b-U152 and G-CTS342 most likely spread from North of the Alps into Italy together, probably in connection with the Bell Beaker expansion."
"According to yfull's estimates, there were only three surviving branches of L497 at 3,200 BC. However, during the first half of the third millennium BC, yfull estimates that L497 underwent a massive expansion and development, with 12 new branches forming in 2,600 BC alone (CTS4803, G727567, Z16770, Y14684, Z16775, Z726, L43, Z1823, Y8903, FGC807, Z1816 and Z6911). All of these branches look West European and are estimated to have arisen during the Bell Beaker expansion period, long before the Urnfield culture and the Celts came into existence. My own estimates, calculated using different methodology, look similar."
"Also according to yfull's estimates, there were only two surviving branches of R1b-L51 at 3,200 BC. However, during the first half of the third millennium BC, yfull estimates that L51 also underwent a massive expansion and development, with scores of new branches forming in Western Europe at the same time that L497 was rapidly expanding and developing in Western Europe. My own estimates also look similar to yfull's in this respect. The early developments of G-L497 and R1b-L51 are both believed to have occurred in South East Central Europe. The best fit for core R1b Bell Beaker mtDNA (across its range) includes a 50% contribution from South East Central European Cucuteni Triploye, which is believed to have been substantially an Early European Farmer G2a population. The Cucuteni Tripolye culture ended in South East Central Europe shortly after 3,500 BC; G-L497 and R1b-L51 in South East Central Europe ended with it. Both L497 and L51 relocated to Western Europe and underwent massive expansions contemporaneously several hundred years later, sharing female DNA in common."
"It looks like Bell Beaker was a hybrid, with components mixed from these two cultures and populations. This mixing was likely to have occurred substantially where the two populations originally co-existed (i.e. in South East Central Europe), and either before or at the time that both relocated contemporaneously from South East Central Europe to Western Europe (i.e. on or before 3,500 BC)." - from eupedia.com/forum
Haplogroup G2a2b2a1a1b is a mutation at L497 with the marker DYS388 = 13. It developed in about 8,800 BC, per Yfull [Scaled Innovation agrees]. Where it developed is an open issue. Most claim a European origin and, indeed, it is today almost entirely a European clade. Its origin may be anywhere from the Balkans, to northern Italy, to central Germany. While I've addressed studies that claim northern Germany, above, I suspect the northern Balkans is more likely. It is most common in southwestern Germany, eastern Switzerland, and western Austria. There are a few samples from North Africa and western Russia north of the Caucasus, but none in the Caucasus itself. This is the most common subgroup of G in Europe, comprising about half of those and 2% of the total population. An unusually large number of SNP's (single nucleotide polymorphism's) are shared by all L497 men,
"suggesting that the L497 group existed as a small group for perhaps a millennium or more before a big expansion began" - per Ray BanksL497 comprises 80% of the G-type samples in the Tyrol (where 10% of the total population are G-types), 75% in Switzerland, 60% in Spain, France and Germany, 52% in England, and 44% in the Netherlands and Italy. By the way, the highest frequencies of L497 in Austria were in the Paznaun Valley, the Upper Inn Valley, the Oetz Valley and the Ziller Valley. Note that Sardinia, which has a large G-type population, has the lowest percentage of L497 recorded in Europe, most of their G-type inhabitants having descended from earlier, Neolithic types.
L497 men in the British Isles often have men from Switzerland and southwest Germany as their nearest continental matches. The separation of the English L497-types from their Swiss and German cousins is calculated, based on the rate of mutation in marker values, to have occurred in the period of the Roman Empire.
". . . but L497 samples outside of Europe continue to be almost unheard of. We were particularly interested last year in a sample from the island of Lemnos off the western coast of Turkey. This island has known connections to the Etruscans. This sample did turn out to be L497+ though it has some very eccentric marker values. And a researcher found a sample with similar marker values from the west coast of Turkey that is also L497 -- the only Turkish sample of this category in his collection. The question remains open whether these men are from an early branch of L497. If so, this could argue for an early movement from the Turkish coast to Europe in conjunction with the Etruscans." - Ray BanksNote however that the most ancient L497 discovered so far was found in Western Ukraine (5310-4860 ybp), making this type a marker for the pre-Indo-Europen language "Old Europe."
Haplogroup G2a2b2a1a1b1 is a mutation at CTS9737 [Z1815]. It probably developed in about 4,700 BC, per Yfull [7,100 BC per Scaled Innovation], probably in southern Germany, near the source of the Danube. Two ancient DNA samples from Baden culture site Budakalasz-Lupa csarda, in Hungary, were typed as CTS9737, circa 3,000 BC. Modern bearers indicate Italian and British Isle ancestry.
Haplogroup G2a2b2a1a1b1a is a mutation at Z1817 [CTS11352, Z759]. It developed around 4,300 BC, per Yfull [4,900 BC per Scaled Innovation], probably in southern Germany or Switzerland, near the source of the Rhine. Ancient DNA from the Viking age had three G2a returns. One could only be tested to G2a, but sample VK39 from Skara-Varnhem [western Sweden], 10th to 12th century AD, was Z1817 (G2a2b2a1a1b1a) and sample VK479 from Gotland-Kopparsvik, 900-1050 AD, was S2808>S23438 (G2a2b2a1a1b1a1a2a1a) [a great-grandchild of Z726, see below].
"Abstract - The Viking maritime expansion from Scandinavia (Denmark, Norway, and Sweden) marks one of the swiftest and most far-flung cultural transformations in global history. During this time (c. 750 to 1050 CE), the Vikings reached most of western Eurasia, Greenland, and North America, and left a cultural legacy that persists till today. To understand the genetic structure and influence of the Viking expansion, we sequenced the genomes of 442 ancient humans from across Europe and Greenland ranging from the Bronze Age (c. 2400 BC) to the early Modern period (c. 1600 CE), with particular emphasis on the Viking Age. We find that the period preceding the Viking Age was accompanied by foreign gene flow into Scandinavia from the south and east: spreading from Denmark and eastern Sweden to the rest of Scandinavia. Despite the close linguistic similarities of modern Scandinavian languages, we observe genetic structure within Scandinavia, suggesting that regional population differences were already present 1,000 years ago. We find evidence for a majority of Danish Viking presence in England, Swedish Viking presence in the Baltic, and Norwegian Viking presence in Ireland, Iceland, and Greenland. Additionally, we see substantial foreign European ancestry entering Scandinavia during the Viking Age. We also find that several of the members of the only archaeologically well-attested Viking expedition were close family members. By comparing Viking Scandinavian genomes with present-day Scandinavian genomes, we find that pigmentation-associated loci have undergone strong population differentiation during the last millennia. Finally, we are able to trace the allele frequency dynamics of positively selected loci with unprecedented detail, including the lactase persistence allele and various alleles associated with the immune response. We conclude that the Viking diaspora was characterized by substantial foreign engagement: distinct Viking populations influenced the genomic makeup of different regions of Europe, while Scandinavia also experienced increased contact with the rest of the continent." - from "Population Genomics of the Viking World" by Ashot Margaryan, et alThe Vikings were not a homogenous ethnicity. These G2a's probably came out of Denmark, i.e. from the Funnelbeaker culture. I should point out that there is a well researched family, the Bures, who are G-L497 (and possibly CTS4803, a subclade of Z726) and come out of Bure, Skelleftea in northeastern Sweden. This family originated in Uppland. There is a large G population (12%) today in Uppsala, Uppland, just north of Stockholm. Overall, however, Sweden is only 2% G.
Still speaking of Vikings and their heirs, Rollo, the conqueror of Normandy, is said per the Icelandic sagas that he was the son of the Jarl of More og Romsdal, in Norway. This is in the fjords, southwest of Trondheim; quite beautiful. The administrator of the FTDNA Norway project has noted that, "in More and Romsdal there are 6/25 hg G [and all L497] and none elsewhere in Norway!"
Haplogroup G2a2b2a1a1b1a1 a mutation at Z727 [Z1816, Z725]. It developed in about 2,500 BC, per Yfull [3,700 BC per Scaled Innovation], probably in southern or central Germany. Those tested positive for this mutation have British and German ancestry. A high percentage of L497 are part of this subgroup.
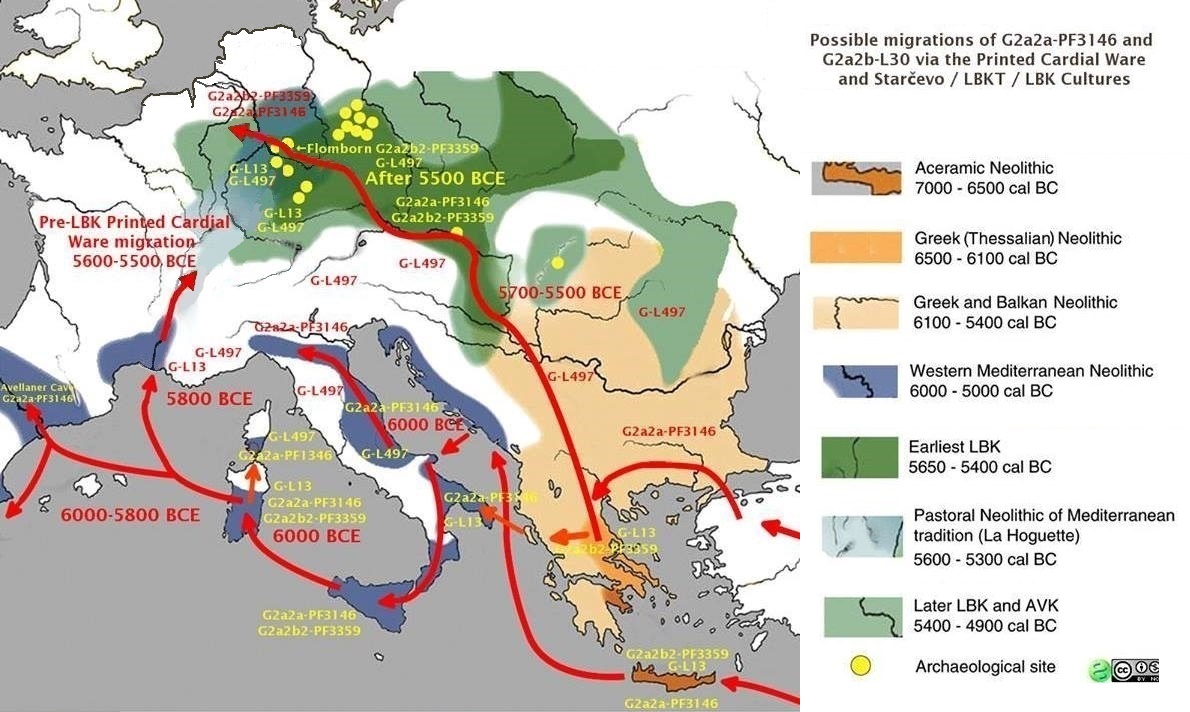
See Haplogroup - M201 for more information about these migations.
Haplogroup G2a2b2a1a1b1a1a is a mutation at CTS2230 [AMM042, Z1823]. It developed in about 2,500 BC, per Yfull [3,500 BC per Scaled Innovation], in western Europe, somewhere between the Rhine and northern France.
Haplogroup G2a2b2a1a1b1a1a2 is a mutation at Z726 [CTS6796]. It probably developed in northwestern Europe in about 2,500 BC, per Yfull [2,700 BC per Scaled Innovation] . . . I'm thinking the Rhine valley, moving towards the sea.
". . . ancestors of [redacted], a retired architect in Cincinnati, Ohio, who is so far the only person in the world to test positive for this [Z726] marker but not for any others further downstream, which could suggest (as Brian Hamman thought in April 2017) that the whereabouts of his ancestors could indicate where the marker arose, about 4,500 years ago (i.e, about 2,500 BC). His known male line ancestry goes back to the Mussgung family of Sollingen near Karlsruhe . . ." - from Adolph and haplogroup G genealogy.Sollingen, near Dusseldorf, is in the Rhine river valley, about 25 miles east of the border with the Netherlands.
Z726 may be tied to the pre-Celtic expansion of the Western Bell Beaker people. See my Celtic Origins page.
". . . during the first half of the third millennium BC, yfull estimates that L497 underwent a massive expansion and development, with 12 new branches forming in 2,600 BC alone [including Z726]. All of these branches look West European and are estimated to have arisen during the Bell Beaker expansion period, long before the Urnfield culture and the Celts came into existence." - from eupedia.comI see modern Y-DNA results for this subclade across Europe, including England, Spain, France, Germany, Denmark, Sweden, and as far east as Ukraine and Russia. Below is a heatmap for Z726, created at Y Heatmap.
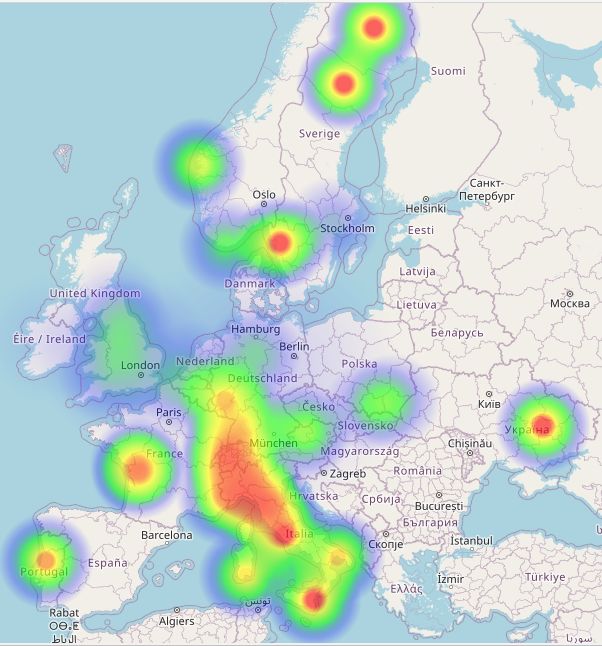
Below is a block tree for the G-Z726 clade, illustraing its relatively short development period; just 5 variants, equating to about 400 years. Note the different development periods of the type's subclades: CTS4803, just 2 variants; Z16775, just 3; Z36217, 46 variants! As you'll read more about below, this means a quick 'evolution' of the first two subclades and a very long bottleneck for the third.
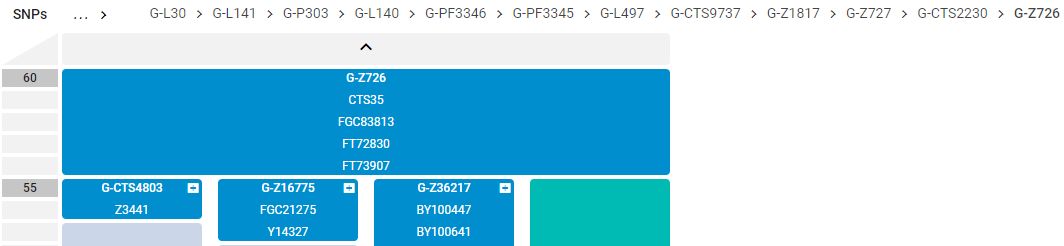
There is a man surnamed Wadsworth who has stated that, "I am English with my paternal ancestry traceable to Yorkshire as far back as the 1700s. My Y-DNA is G2a3b1a [old designation], specifically G-Z726." I think, however, he is part of the G-Z16775 subclade. And, I just found a reference to a Thomas Henry Eason (1825-1911) of Whaplode Drove, Lincolnshire. His descendant, Eric Louis Eason got a DNA test from Living DNA, which yielded G-Z726. Could he be a Heesom whose surname got respelled?
Haplogroup, G2a2b2a1a1b1a1a2c is a mutation at Z36217 [Z36218]. It formed around 2,200 BC, per Yfull [2,150 BC per FTDNA, 1,800 BC per Scaled Innovation]. The Most Recent Common Ancestor was born circa 1150 AD, per FTDNA. That is at the beginning of the naming period when (4) Adam Gernet (c1140) of Heysham became known as Adam de Hessam or de Heysham. So far, all the results are for families from England. The Hissem family, which can trace their origins back to the West Riding, Yorkshire, England and perhaps further back to Lancashire, is part of the Z36217 group. This includes Crispin Heesom, of England, and Steve Hissem, James Olin Hissom and Alan Hissem of America. Justin Heysham of Australia, if he gets a Y-DNA test, may be of this group as well.
Below is a heat map for Z36217 as well as heat maps for the major Z726 subclades, Z16775 and CTS4803. These are derived from Y Heatmap. Note that only Z36217 is an England-only clade. See the Bell Beaker Culture for what I think are the ramifications of this. An administrator at FTDNA writes,
"Currently all the confirmed G-Z36217 men, English, share a patrilineal ancestor who lived less than 1000 years ago, so we do know that at around 1000 AD the branch was probably in UK already, but we don't have great clues regarding possible (pre-)G-Z36217 movements between 4200 and 1000 years ago."The origins of Z16775 and CTS4803 are in the upper-Rhine to northern Italy, while that for Z36217 is in the middle of England. Also of note, Z16775 and CTS4803 have 57 and 174 subclades, respectively, Z36217 has just 5. I suppose that means those types are more common and their progenitors more prolific, and our type relatively rare. You know, like a fine wine.
The map below, from Scaled Innovation, shows a possible path for haplogroup G2a to get to Britain, in this case down the Rhine, through the Rhenish Bell Beakers' homeland, evolving to G-Z726, and finally to Britain, where it evolved into G-Z36217.
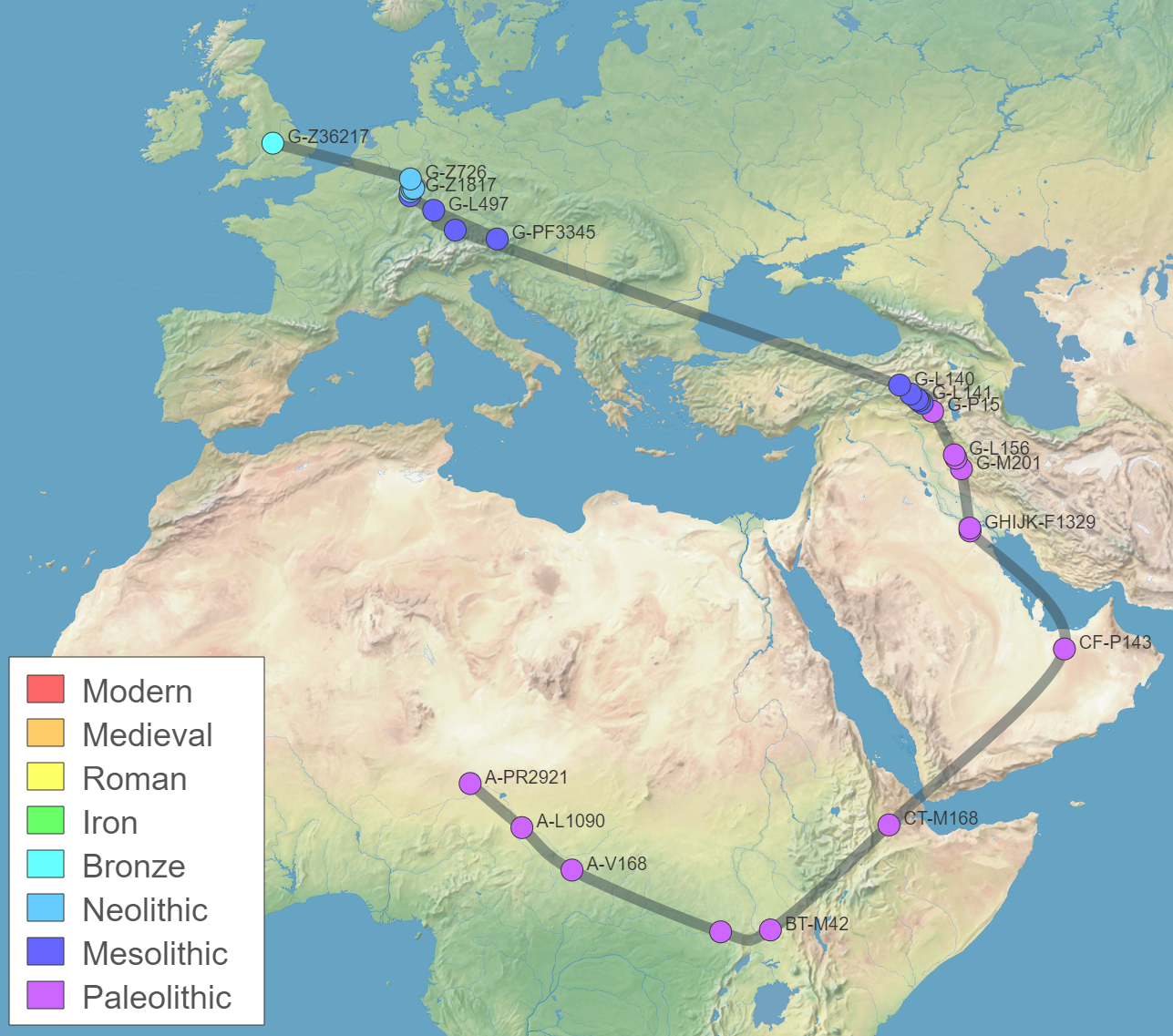
Another view of this path is via Family Tree DNA, below.
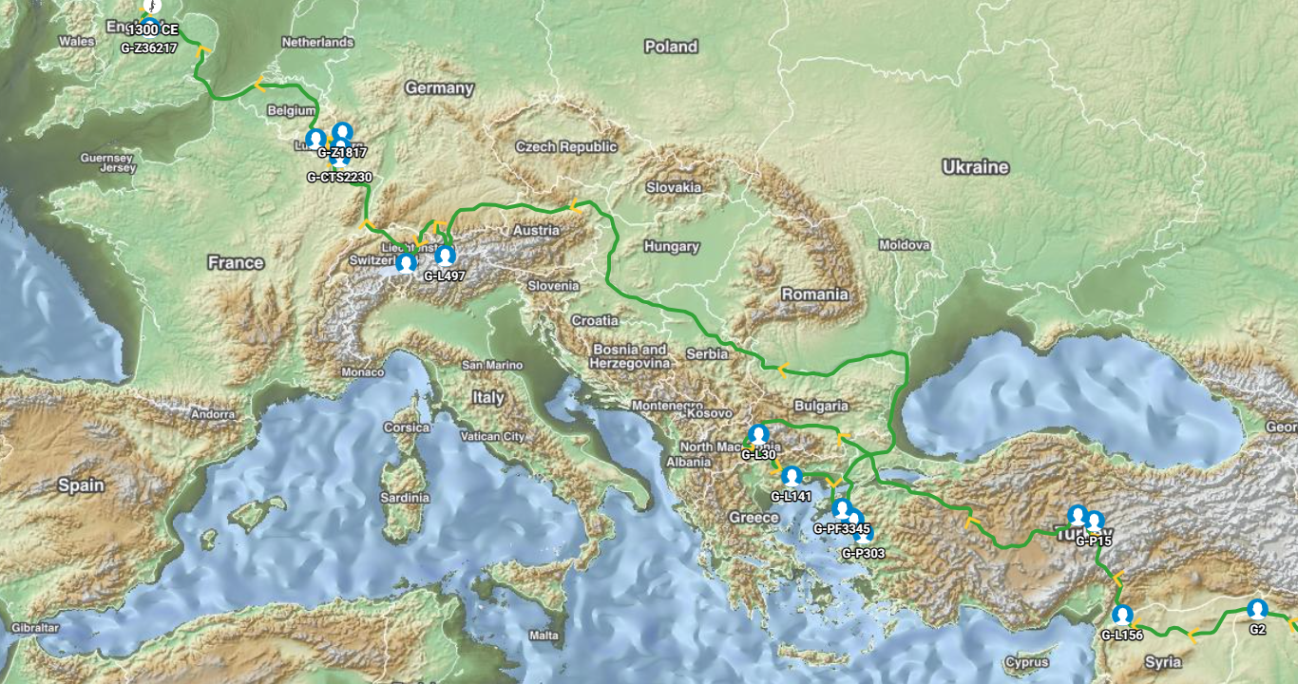
One measure of a clade is its number of equivalent SNPs (also known as variants or mutations) within its branch. Z36217 is unique in having a very long branch length at 46 variants, and in having only two sub-clades, and another three below those. Its sister clades have only a handfull of branch variants and very many sub-clades.
| Branch Length in Phylogenetic Trees
Branch length is a measure of divergence or genetic change in a lineage. The units of branch length are the number of changes or mutations. These are not time estimates, but are considered a good proxy for time. Per FTDNA, "branch lengths can represent evolutionary time, calculated as number of mutations."As a general rule, each mutation (an SNP or variant generation) equates to 80 to 140 years. In the example below, G-FT115823 would have a block length of 3, G-FT115839 would have a block length of 6, and G-FT248728 a block length of 4. There are 11 SNPs identified between, roughly, today and the formation of FT115823. This equates to about 1100 years. I say 'roughly, today' because I think we should actually count from the birth year of the subject.    The longer the length of the branch, the more genetic change has occurred without sub-clades forming. A long branch length implies a bottle-neck of some sort. G-Z36217, which supposedly emerged in about 2,200 BC, has a branch length of 46 SNP mutations/variants and its next subclades did not emerge for 3,300 years. That is a very 'skinny' line of descent with no 'brothers' creating their own lines of descent.
Z36217's sister clades, CTS4803 and Z16775, have 2 and 3 branch variants, respectively, and an extensive list of subclades. Z16775 has a TMRCA just one hundred years after the SNPs emergence in 2,200 BC. CTS4803 has a TMRCA that is the same as its time of emergence. These differences must be significant. The following is a list of our ancestor SNPs, going backwards in time, their branch length or number of variants, approximate date of emergence, time to the Most Recent Common Ancestor, and years elapsed from previous clade to current version/years per variant. Dates are from Yfull; other sites have different dates. I'm not sure how valid TMRCA is considering that it uniformly matches the formation date of the child clade.
**One of the longest block lengths ***The G haplogroup parent group has been identified as suffering a long bottleneck before its first subclades, G1 and G2, emerged. Assuming 80 years per variant, that's over 25,000 years! That would mean that of the G-types approximately 45,000 years of existence, in 55% of that time it was confined to just the parent clade. |
So it appears that our Z36217 line of the family suffered a bottleneck from 2,000 BC to 1,100 AD as evidenced by its branch length. What might cause this?
The reduction in the size or a brake on the natural increase of a population may be caused by natural events, famine, disease, or war. Carried to an extreme this becomes not a bottleneck, but an extinction. I would guess that in our case this brake on our development was simply the result of hard times. Some new clades probably did emerge from Z36217, but they did not survive. Finally, around 1100 AD (the Z36217 TMRCA), new lines of descent were able to prosper. What changed about this time? Well, the last successful invasion of Britain had occurred in 1066 and been followed by the harrying of the north.
| Harrying of the North
"The Harrying of the North was a number of campaigns waged by William the Conqueror in the winter of 1069-70 to subjugate northern England, where the presence of the last Wessex claimant, Edgar Atheling, had encouraged Anglo-Danish rebellions. William paid the Danes to go home, but the remaining rebels refused to meet him in battle, and he decided to starve them out by laying waste to the northern shires using scorched earth tactics, especially in the city of York, before relieving the English aristocracy of their positions, and installing Norman aristocrats throughout the region. Contemporary chronicles vividly record the savagery of the campaign, the huge scale of the destruction and the widespread famine caused by looting, burning and slaughtering. Some present-day scholars have labelled the campaigns a genocide, although others doubt whether William could have assembled enough troops to inflict so much damage and have suggested that the records may have been exaggerated or misinterpreted. Records from the Domesday Book show that 75% of the population died or never returned. "The King stopped at nothing to hunt his enemies. He cut down many people and destroyed homes and land. Nowhere else had he shown such cruelty. This made a real change. To his shame, William made no effort to control his fury, punishing the innocent with the guilty. He ordered that crops and herds, tools and food be burned to ashes. More than 100,000 people perished of starvation. I have often praised William in this book, but I can say nothing good about this brutal slaughter. God will punish him." - Orderic Vitalis"The land was ravaged on either side of William's route north from the River Aire. His army destroyed crops and settlements and forced rebels into hiding. In the New Year of 1070 he split his army into smaller units and sent them out to burn, loot, and terrify. Florence of Worcester said that from the Humber to the Tees, William's men burnt whole villages and slaughtered the inhabitants. Food stores and livestock were destroyed so that anyone surviving the initial massacre would succumb to starvation over the winter. The survivors were reduced to cannibalism." - from Wikipedia. |
So, once these ugly times had ended, our family had peace enough to prosper and engender, no matter how slowly, new branches to the family.
G-Z36217 has just two sub-clades at this time, FT115823 and Z38868, the latter being the Heesom/Hissem family terminal clade.
Haplogroup G2a2b2a1a1b1a1a2c1 is a mutation at FT115823. This is for the Whittington family.
Haplogroup G2a2b2a1a1b1a1a2c1a (I think) is a mutation at FT115839 for an descendant of James Whttington, though I don't see it in ISOGG. A Whittington family researcher says,
"The person who tested was Joseph Allen Whitenton. James Whittington born 1776 in Johnston County, NC is our common ancestor. My line follows his son, George A. Whitenton and Joseph is a descendant of Solomon Whitenton."
Haplogroup G2a2b2a1a1b1a1a2c1a1 is a mutation at FT115715. It was found for Mike and Walter Whitenton. We share 101 of 111 STRs. It formed approximately 820 years ago, per Yfull, or 1132 AD.
Haplogroup G2a2b2a1a1b1a1a2c1a2 (I think this will change to G2a2b2a1a1b1a1a2c1b based on the FTDNA Block Tree) is a mutation at FT248728. It was found for Karl, Tom and Jeremy Whittington, all matches for me at FTDNA; we share 101 of 111 STRs. I received the following from Diane Whittington Bollert,
"I just wanted to let you know how much I enjoy reading about The Hissem-Montague Family. As far as I know I am not a Hissem descendant, but you never know. My dad, Walter Whitenton (desc of Richard Whittington b abt 1710 in NC) tested at FTdna and is confirmed G-Z36217. I have not found a connection to the Whittington McGlamery line or to Brian Fletcher who shares the same terminal snp. I still have no proof where my family came from in England, but family legend suggests they were from Nottinghamshire. I recently took the plunge and ordered the BigY700 test and the results are due back this month. Hopefully my dad's results will add another piece to the puzzle and we will all benefit from it. My dad's kit number in B149098." - from Diane Bollert nee WhitentonThere is a 98.01% possibility that Karl and I share a common ancestor at generation 24; 91.85% at generation 20; and 74.02% at generation 16. I suspect that Gerald McGlamery, whose father line is that of Whittington, is also part of this sub-clade. Gerald is also a match for me on FTDNA. Compare this with Brian Fletcher, below, who much more closely related.
Haplogroup G2a2b2a1a1b1a1a2c2 is a mutation at Z38868 [BY135198, BY77286, Z38867]. It is the other subclade of Z36217. ISOGG lists this as an Investigational Item, not yet confirmed. It formed about 900 years ago, per Yfull, or 1100 AD [1150 AD per FTDNA, 1400 AD per Scaled Innovation]. This was during the first century of Norman rule in England, when Roger Gernet was Chief Forester of Lancashire and Lord of the village of Hessam (Heysham). This was also an era when surnames began to be more common.
G-Z38868 is the Hissem family terminal clade, at least so far. Brian L. Fletcher is a match for me on FTDNA. An administrator at Family Tree DNA writes,
"Now, based on the current data and YFull methodology, I believe this new G-Z38868/G-BY135198's age (TMRCA - Time to Most Recent Common Ancestor) would be estimated in 450-350 ybp (years before present) or so, i.e., the most recent common patrilineal ancestor Mr. Fletcher and yourself share would be from about 1600 AD. Keep in mind that this estimation can slightly change occasionally, with further data; it's just a reference."The following is how I think the administrator made this estimate. The block length of Z38868 is 4 mutations (Z38868, BY135198, BY77286, FT250916). Using a time for each mutation of 100 years yields 400 years ago or 1600 AD. That is the approximate birth year of the MRCA for the English and American branches of our family, George Heesom of Crofton, Yorkshire. FTDNA predicts a TMRCA of 1550, or Raphe Heesome of Barnsley, Yorkshire.
However, the TiP calculator on Family Tree DNA predicts that Brian L Fletcher and I share a common ancestor 19 generations ago at a 96% level. I use this percentage because I and Alan Hissem Jr. share a common ancestor 7 generations ago (I have good paper trail to support this) and the FTDNA predicts this accurately at the 95% level. 19 generations is approximately 570 years (30 years per generation), or 1382.
Brian and I share 809,180 variants (FTDNA calls SNPs variants) and have seven non-matching variants. Three of these non-matching variants he has (16964617, 4800349, 6279634) and I do not. Four of these I have (3242901, 7168342, 17164318, 19639921) and he does not; they are my private variants. Private variants are SNP mutations that have occurred in the line being tested, but have not yet been named. Occurrences of private variants in multiple testers allow the SNP to be named and placed on the haplotree. Since Brian doesn't share my private variants, nor I his, then we are part of diffent subclades of Z38868. This makes sense if the predicted MRCA, above, is correct since my ancestor left England no later than 1686, not long enough for a new mutation to have formed. Neither of these subclades of Z38868 can be named until another tester comes along and matches one of us. For me, this will have to be someone in the American line of the family. The following is the Block Tree for the subclades of G-Z36217.
Since Z38868 has a block length of 4, for an emergence circa 1600, and I have 4 private mutations or variants, to bring us up to the present, does that mean that my latest mutation, which would describe my terminal clade, must have emerged very recently? If so, then Brian Fletcher, with 3 private variants, must have a terminal clade that is at least 80 years older than my own.
Below is the Block Tree for G-Z36217, including all the sub-clades so far identified. Note the large number of equivalent variants. G-36217 emerged in about 2200 BC and G-Z38868 emerged in about 1100 AD.
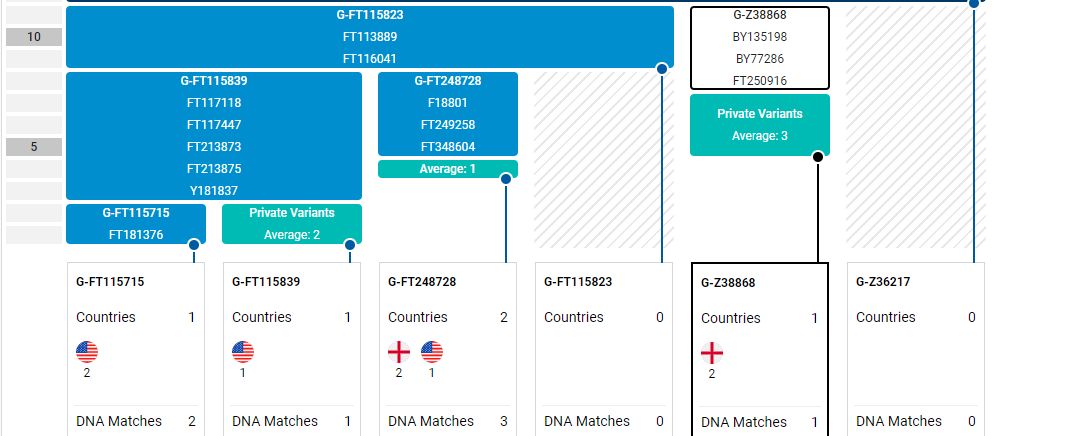
The Y-67 and Y-111 tests identify STRs, Short Tandem Repeats, which mutate more rapidly than SNPs. I and the Heesoms of Yorkshire vary by just 3 STRs out of 67. Brian Flether and the Heesoms of Yorkshire also vary by 3 out of 67, though not for all the same STRs as myself. Brian and I vary by 4 out of 67, and 7 out of 111.
The following are also predicted to be Z36217 and therefore a match for me.
- The John Clair family, who trace their origins to William T. Clear.
- The Scott family, who trace their origins to a William Scott, who married in 1639 in North Molton, Devon.
The new block diagram is as follows. Newly identified clades include Y183439, FT411024, F15541, FT409494, and FT411158. Alan Hissem, Crispin Heesom, and I have a most recent common ancestor of Thomas Hesom, born circa 1720 in Trenton, New Jersey.
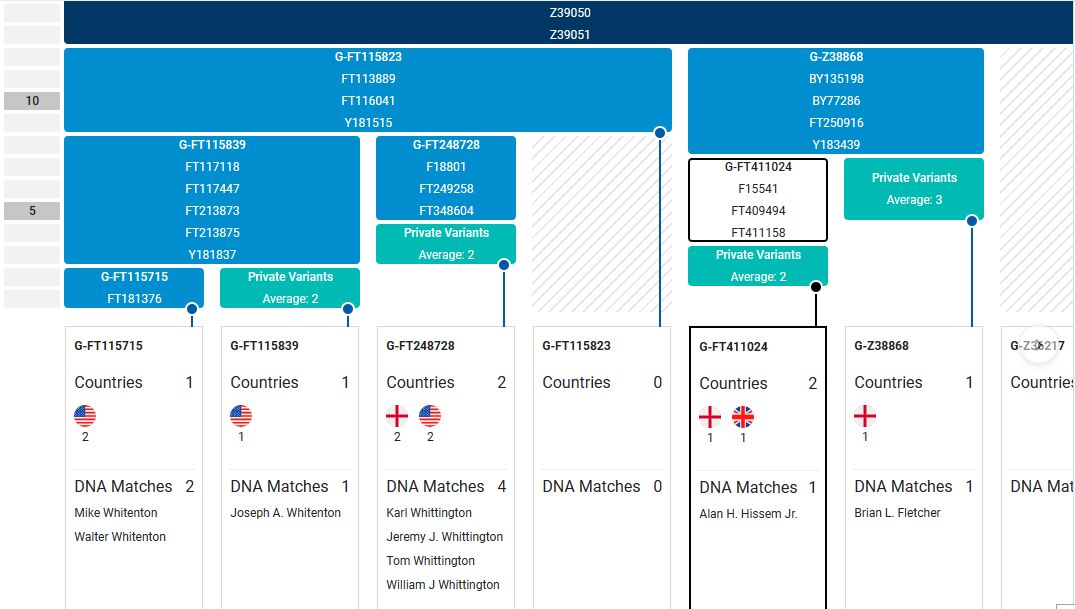
Below is the haplogroup G2a phylogenetic tree.
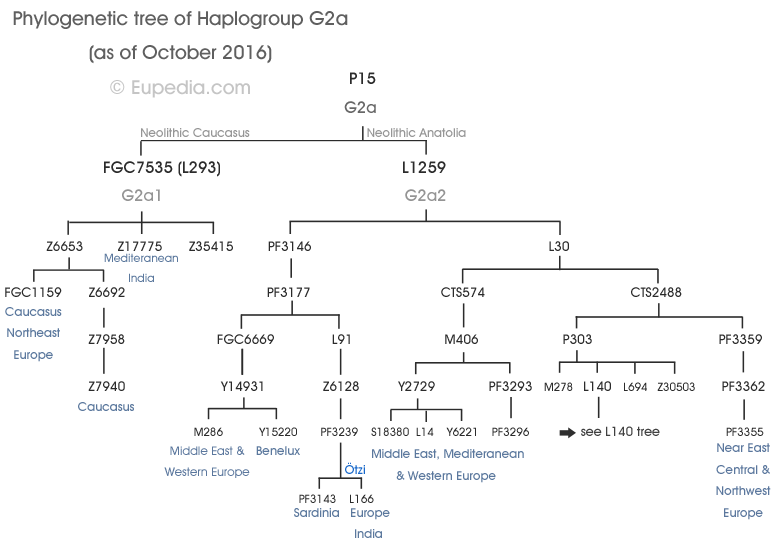
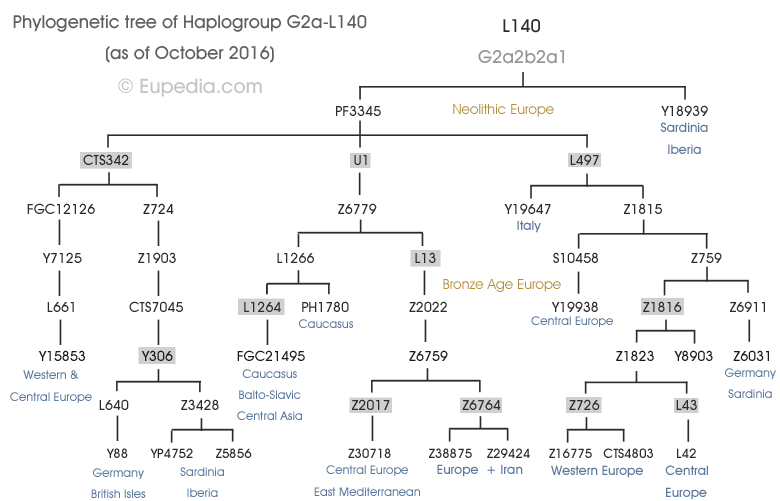
The charts below break down the distribution of the G haplotype by its major subclades. They are from Eupedia.com.
G-M201 defines the basic haplogroup and so includes all G-types.
"The general frequency pattern of hg G overall shows that the spread of hg G extends over an area from southern Europe to the Near/Middle East and the Caucasus, but then decreases rapidly toward southern and Central Asia." - "Distinguishing the co-ancestries of haplogroup G Y-chromosomes in the populations of Europe and the Caucasus" by Siiri Rootsi, et alWhile the concentration in the Caucasus stands out, note the vector across Turkey into western Anatolia and the hotspots in northern Greece.
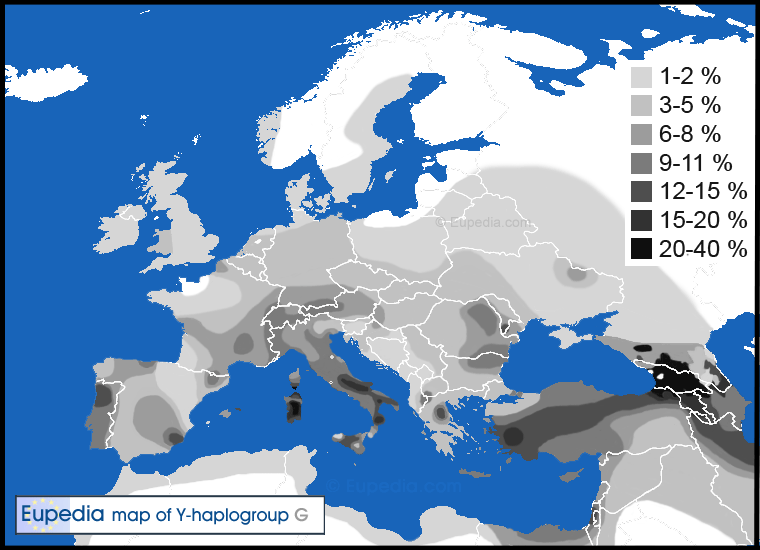
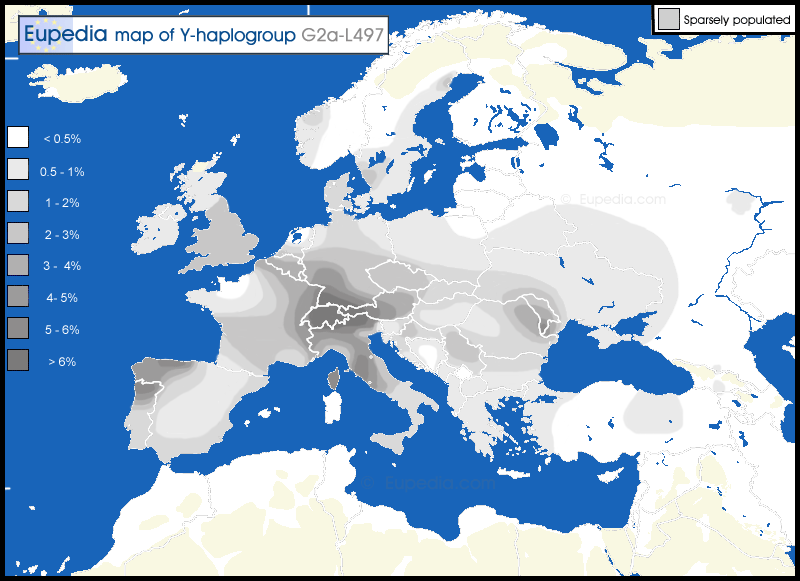
By the way, the Y-DNA of the Hysom family of Dorset has been tested. The sample I saw did not have a haplogroup identified, so I used a haplogroup predictor. It yielded a prediction of E3b-V13 [E3b1a2] at a 68% level, or E3b-M78 [E3b1a] at 31%. So, they are not relatives of the Heesom family, despite the phonetic similarity of their surname. No Y-DNA testing of the Heysham family of Lancaster, our presumed ancestors, has been done to my knowledge.
Another recent find was the Y-DNA test of the Garnett famiy, possible descendents of the Norman Gernets. However a trip through the Haplogroup predictor showed them to be R1b, the standard English type, not G2a.
(27) Alan Howard Hissem Jr. (c1956), Genetic Distance: 2 (of the Thomas Hissom branch of the American line of family)
Brian Fletcher, Genetic Distance: 9, share 102 of 111 markers, 6 of 592 STR differences, Z726>Z36217>Z38868.
"A 102/111 match indicates a distant cousinship with a chance of a genealogical relationship. Over half of matches at this level are related as 13th cousins or closer. Most matches at this level are related as or more recently than 20th cousins. The connections here can be highly informative for relationships with historic groups and events. If there is a tradition of a recent genealogical relationship, the best way to confirm it is to test additional family lines. By testing additional family lines, you can find the person in between who is a closer match to each of the others tested. This 'in betweener' is essential for you to find as their match proves the connection between the more distant matches." - from FTDNASo, a 13th to 20th cousin perhaps. A 13th cousin shares an ancestor 14 generations ago. My paper trail extends to an ancestor 10 generations past, born in about 1600. That's about 350 years prior to my own birth, so about 35 years per generation. Pushing that back 4 more generations to the 14th, at the same rate, makes 1460. Another 7 for the 21st makes 1215. So Brian and I share a common ancestor born between 1215 and 1460, per our genetic distance of 9. However, I will point out that I have, per my paper trail, a 9th cousin living in England (Crispin Heesom). According to TiP, an 80% confidence level yields a common ancestor with him 10 generations back (probably George Hesom, born c1600). Using the same 80% confidence level for Brian yields a common ancestor at generation 14, that is 1460 (I have a number of Hesams, Heshams, Hesehams and Heyshams born in Lancaster and York c1400-c1450). This was a time of great turmoil in England during the internecine struggles of the War of the Roses (1455-1487) that followed England's final defeat by France in the Hundred Year's War (1337-1453).
Karl Whittington, Genetic Distance: 10, 9 of 562 STR differences, Z726>Z36217>FT115823>Z248728. The TiP calculator yields a common ancestor at generation 16 at approximately 80% confidence (actually 74%), i.e. about 1390.
Jeremy Whittington, Genetic Distance: 10, 11 of 567 STR differences, Z726>Z36217>FT115823>Z248728. The TiP calculator yields a common ancestor at generation 16 at 74% confidence, i.e. about 1390.
(27) Crispin David Edwin Heesom (c1950), Genetic Distance: 3 (of the East Yorkshire line of the family)
Jeremy J. Whittington, Genetic Distance: 5
Tom Whittington, Genetic Distance: 6, 11 of 569 STR differences, Z726>Z36217>FT115823>Z248728.
Gerald Garris McGlamery Jr., Genetic Distance: 6. Gerald wrote me,
"Thanks for reaching out. I just saw my results yesterday for the first time, and I'm still trying to decide what they tell me. Several years ago, I wanted to take a Y-DNA test to better understand the origins of the McGlamery name for my own site (http://www.mcglamery.com/blog/mcglamery-genealogy/), but I procrastinated. Then, a couple of years ago, a cousin came across a memoir from where she grew up in Wilkes County, NC, that in a footnote stated that my great-great-grandfather, Murphy Augustus McGlamery (1847-1921), was, in fact, the son of Allen Whittington (1801-1899). Since Murphy was born a couple of months after his reported father, David McGlamery (1800-1846), died, I certainly understood that Allen Whittington could well have been the biological father. I recently retired and finally got around to taking the Y-67 test, and sure enough, there are a couple of Whittingtons that showed up in the match list, though at the 25- and 37-marker level. There are no McGlamerys. Consequently, my Y-DNA appears to route back through the Whittingtons of Wilkes County, NC.
I already have two lines of Whittingtons in other branches of my family, and one of them is Allen. I also descend from Alexander Greenberry Whittington (1811-1890), Allen's brother. All lines meet at their father, Leonard Whittington (1770-1856). Leonard's father was William Whittington (1736-1807), who came to Wilkes County from Virginia. Some genealogists place his birth in Anne Arundel County, MD. Until now, I haven't really spent any time trying to understand William Whittington, but I guess my interest level just went up."
Joseph Allen Whitenton, Genertic Distance: 7, 12 of 569 STR differences, Z726>Z36217>FT115823>FT115839.
John Patrick Clair, Genetic Distance: 7
William J. Whittington, Genetic Distance: 7
Matthew Dayton Chaiser, Genetice Distance: 7 (paternal line under different surnames). Born 14 May 1955 to Robert L. Chaiser and Rita J. Blais. Robert L[eroy] Chaiser born 19 December 1917 to Edwin J. Chaiser and Mae Cnnstance Burkman, died 22 November 1998. Edwin John Chaiser born on 11 August 1885 in Illinois the son of John and Matilda Chaiser, died 22 February 1972. John J. Chaiser, the husband of Mathilda L. Astrand [Ostrand], was born in Sweden, as was Mathilda, he on 1 August 1857, she in 1859. John and Matilda emigrated in 1880. John died on 11 March 1940, Matilda in 1945. The Chaiser name appears to have been from Uppsala, Sweden. John J. Chaiser may be the son of Jonas Peter Magnuson and Christiana "Pettersdotter." She was born on 1 July 1817.
(26) James Olin Hissom (1923), Genetic Distance: 3 (of the David Heysham branch of the American line of the family)
Andrew James Peel, Genertic Distance: 1, 28 of 537 STR differences, L43>L42>>Y11074>Z45255>Z45259. Of Ireland.
Thomas J. Allaire, Genertic Distance: 2, 23 of 557 STR differences, Z726>CTS4803>Z6150>Z30771>BY28016. Of France.
Kevin Stickle, Genertic Distance: 2, 29 of 584 STR differences, L43>L42>Y11074>Y11076>CTS7357>Z44718>Z45417. Of Pfalz, Germany.
Christian Freytag, Genertic Distance: 2, 30 of 603 STR differences, Z726>CTS4803>S2808>Z42514>Z31329. Of Germany.
 While we tend to talk about Anatolia as being analgous to today's Turkey, it anciently was considered to comprise only that part which is a penisula of Asia. It was for that reason known as Asia Minor, "lesser asia," in classical times. It was bounded by the Black Sea to the north, the Mediterranean Sea to the south, and the Aegean Sea and Sea of Marmara to the west. In the east it extended to the Armenian Highlands, also known as Armenia Major, and bordered the Euphrates river before that river bends to the southeast to enter ancient Mesopotamia.
While we tend to talk about Anatolia as being analgous to today's Turkey, it anciently was considered to comprise only that part which is a penisula of Asia. It was for that reason known as Asia Minor, "lesser asia," in classical times. It was bounded by the Black Sea to the north, the Mediterranean Sea to the south, and the Aegean Sea and Sea of Marmara to the west. In the east it extended to the Armenian Highlands, also known as Armenia Major, and bordered the Euphrates river before that river bends to the southeast to enter ancient Mesopotamia. The Neolithic, or New Stone Age, began when man invented farming and so varies in time around the world. In the Middle East, where it is believed farming first originated, it began in about 10,000 BC. Early Neolithic farming was limited to a narrow range of plants, both wild and domesticated, which included einkorn wheat, millet and spelt, and to the keeping of dogs, sheep and goats. By about 6,900-6,400 BC it included domesticated cattle and pigs and the establishment of permanently or seasonally inhabited settlements. The period is generally divided into four eras,
The Neolithic, or New Stone Age, began when man invented farming and so varies in time around the world. In the Middle East, where it is believed farming first originated, it began in about 10,000 BC. Early Neolithic farming was limited to a narrow range of plants, both wild and domesticated, which included einkorn wheat, millet and spelt, and to the keeping of dogs, sheep and goats. By about 6,900-6,400 BC it included domesticated cattle and pigs and the establishment of permanently or seasonally inhabited settlements. The period is generally divided into four eras,
 A Neolithic settlement in southeastern Turkey, at the foot of the Taurus mountains. It prospered from circa 8,630 to 6,800 BC, a period that covers the Pre-Pottery Neolithic A & B (PPNA & PPNB) and Pottery Neolithic (PN). It lies near the Bogazcay river, a tributary of the upper Tigris River. Archeological finds include farming tools that suggest both crops and animal husbandry as well as domestication of the dog. Religion is represented by figurines of Cybele, a mother goddess.
A Neolithic settlement in southeastern Turkey, at the foot of the Taurus mountains. It prospered from circa 8,630 to 6,800 BC, a period that covers the Pre-Pottery Neolithic A & B (PPNA & PPNB) and Pottery Neolithic (PN). It lies near the Bogazcay river, a tributary of the upper Tigris River. Archeological finds include farming tools that suggest both crops and animal husbandry as well as domestication of the dog. Religion is represented by figurines of Cybele, a mother goddess.  However, after long centuries of warming earth's temperature took a sudden and precipitous drop. This ushered in a 1000-year cold snap called the Younger Dryas, between 12,000 and 13,000 years ago, and had a profound effect on man's history. When the cold temperatures of the Younger Dryas intervened the wild cereals were crowded out by scrub that could better survive the new, colder, drier environment. It is thought that farming began as people cleared away the scrub, creating beds in which the wild cereal they had become dependent upon could prosper. Inspired by this they began to sow the wild seed themselves and, eventually, to domesticate and improve the plants. The success of their efforts resulted in the Neolithic farming revolution. With a more secure food source, a mankind of limited numbers and precarious existence was transformed into a burgeoning mankind of towns and cities that could manipulate their natural environment and free others to create new technologies in other fields.
However, after long centuries of warming earth's temperature took a sudden and precipitous drop. This ushered in a 1000-year cold snap called the Younger Dryas, between 12,000 and 13,000 years ago, and had a profound effect on man's history. When the cold temperatures of the Younger Dryas intervened the wild cereals were crowded out by scrub that could better survive the new, colder, drier environment. It is thought that farming began as people cleared away the scrub, creating beds in which the wild cereal they had become dependent upon could prosper. Inspired by this they began to sow the wild seed themselves and, eventually, to domesticate and improve the plants. The success of their efforts resulted in the Neolithic farming revolution. With a more secure food source, a mankind of limited numbers and precarious existence was transformed into a burgeoning mankind of towns and cities that could manipulate their natural environment and free others to create new technologies in other fields. Mankind has, through selective breeding, influenced the development of both plants and animals. These domesticated breeds were selected, sometimes inadvertantly, for desirable traits. For example, wild wheat "shatters and falls to the ground to reseed itself when ripe, but domesticated wheat stays on the stem for easier harvesting. This change was possible because of a random mutation in the wild populations at the beginning of wheat's cultivation. Wheat with this mutation was harvested more frequently and became the seed for the next crop. Therefore, without realizing, early farmers selected for this mutation. The result is domesticated wheat, which relies on farmers for its reproduction and dissemination." - from Wikipedia.
Mankind has, through selective breeding, influenced the development of both plants and animals. These domesticated breeds were selected, sometimes inadvertantly, for desirable traits. For example, wild wheat "shatters and falls to the ground to reseed itself when ripe, but domesticated wheat stays on the stem for easier harvesting. This change was possible because of a random mutation in the wild populations at the beginning of wheat's cultivation. Wheat with this mutation was harvested more frequently and became the seed for the next crop. Therefore, without realizing, early farmers selected for this mutation. The result is domesticated wheat, which relies on farmers for its reproduction and dissemination." - from Wikipedia. The horses of the Ice Age were hunted by early modern humans for meat in Europe, across the Eurasian steppes, and into North America. Cave paintings in Europe indicate what these horses looked like. They had large heads, thick necks, stiff upright manes, relatively short, stout legs, and a dun coloration. The horse was first domesticated in the Eurasian Steppes in approximately 3,500 BC. Apparently this occurred among the Botai people of Akmola Province in Kazakhstan. They rode horseback to hunt wild horses, there being no other large prey animals in that region. They also milked their horses. Under domestication the horse evolved into its current larger, longer legged version.
The horses of the Ice Age were hunted by early modern humans for meat in Europe, across the Eurasian steppes, and into North America. Cave paintings in Europe indicate what these horses looked like. They had large heads, thick necks, stiff upright manes, relatively short, stout legs, and a dun coloration. The horse was first domesticated in the Eurasian Steppes in approximately 3,500 BC. Apparently this occurred among the Botai people of Akmola Province in Kazakhstan. They rode horseback to hunt wild horses, there being no other large prey animals in that region. They also milked their horses. Under domestication the horse evolved into its current larger, longer legged version.  The story of the very early city of Gobekli Tepe, circa 9,000 BC, in southeast Turkey on a tributary of the Tigris river, upends the discussion above about the origins of settled life. Here it appears that the settling down, which apparently occurred for religious purposes (the explanation for anything we don't understand), led to the later development of an agricultural economy. It is here that the earliest stone buildings are found.
The story of the very early city of Gobekli Tepe, circa 9,000 BC, in southeast Turkey on a tributary of the Tigris river, upends the discussion above about the origins of settled life. Here it appears that the settling down, which apparently occurred for religious purposes (the explanation for anything we don't understand), led to the later development of an agricultural economy. It is here that the earliest stone buildings are found.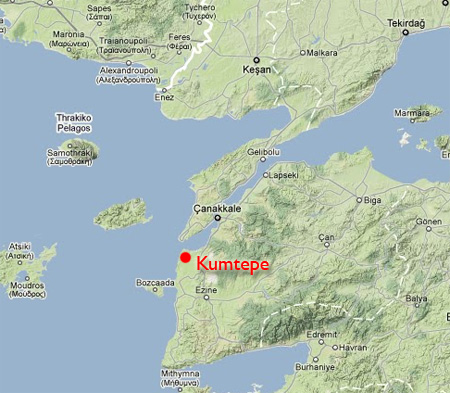 "Kumtepe is the oldest permanent settlement in the Troas, the region in northwestern Anatolia, where later Troy was built. Kumtepe has four layers, Kumtepe IA, IB, IC and II. The last two have been largely disturbed in the twentieth century. The remaining and relatively undisturbed IA and IB are of special interest to the archaeologists, because these are older than other settlements in the region.
"Kumtepe is the oldest permanent settlement in the Troas, the region in northwestern Anatolia, where later Troy was built. Kumtepe has four layers, Kumtepe IA, IB, IC and II. The last two have been largely disturbed in the twentieth century. The remaining and relatively undisturbed IA and IB are of special interest to the archaeologists, because these are older than other settlements in the region. The earliest Neolithic Farmers in Europe arrived from Anatolia and settled first in Greece, around 6,500 BC, roughly coincident with their settlement of western Turkey. They may have arrived via a still existent land bridge at the Bosphorus, or used primitive boats to paddle across the Aegean, moving from island to island, never losing site of land.
The earliest Neolithic Farmers in Europe arrived from Anatolia and settled first in Greece, around 6,500 BC, roughly coincident with their settlement of western Turkey. They may have arrived via a still existent land bridge at the Bosphorus, or used primitive boats to paddle across the Aegean, moving from island to island, never losing site of land. These were three related Neolithic cultures, successors to the early Vinca culture of southeastern Europe, see map below. They were located in Serbia, eastern Hungary and Romania, respectively. Their pottery was usually coarse, but finer fluted and painted vessels later emerged. - from Wikipedia.
These were three related Neolithic cultures, successors to the early Vinca culture of southeastern Europe, see map below. They were located in Serbia, eastern Hungary and Romania, respectively. Their pottery was usually coarse, but finer fluted and painted vessels later emerged. - from Wikipedia.
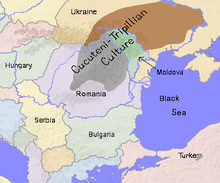 The late Neolithic Cucuteni-Trypillian culture, from Western Ukraine, Moldova, and Romania, produced an enormous farming population. They lived alongside and just to the west of the Yamnaya culture steppe nomads, who were type R1a/b. When the climate suddenly got much warmer and drier around 3,500 BC the Cucuteni-Trypillian sites were abandoned because the region became too dry to support such a large population. The Yamnaya began a westward migration soon after, supplanting the Cucuteni-Trypillians.
The late Neolithic Cucuteni-Trypillian culture, from Western Ukraine, Moldova, and Romania, produced an enormous farming population. They lived alongside and just to the west of the Yamnaya culture steppe nomads, who were type R1a/b. When the climate suddenly got much warmer and drier around 3,500 BC the Cucuteni-Trypillian sites were abandoned because the region became too dry to support such a large population. The Yamnaya began a westward migration soon after, supplanting the Cucuteni-Trypillians. The people who produced Linear Pottery were the first true farmers in central Europe. Their crops included wheat, crab apples, peas, lentils, flax, linseed, poppies and barley. Domestic animals included cattle, sheep and goats. They lived in small villages alongside waterways and built large longhouses with rectangular plans.
The people who produced Linear Pottery were the first true farmers in central Europe. Their crops included wheat, crab apples, peas, lentils, flax, linseed, poppies and barley. Domestic animals included cattle, sheep and goats. They lived in small villages alongside waterways and built large longhouses with rectangular plans.  This culture, 4,100-2,650 BC, represented a merger of Linear Pottery culture derived agricultural and Mesolithic hunter-gatherer lifestyles. It stretched across the north European plain from the Netherlands to Poland, and into Scandinavia. The economy included small scale farming of wheat and barley, but was dominated by the raising of sheep, cattle, pigs and goats. The people lived in wooden longhouses inside fortified hill-top villages. During their period of dominance megalithic tombs and passage graves of the Atlantic Megalithic culture appeared, especially in the Netherlands.
This culture, 4,100-2,650 BC, represented a merger of Linear Pottery culture derived agricultural and Mesolithic hunter-gatherer lifestyles. It stretched across the north European plain from the Netherlands to Poland, and into Scandinavia. The economy included small scale farming of wheat and barley, but was dominated by the raising of sheep, cattle, pigs and goats. The people lived in wooden longhouses inside fortified hill-top villages. During their period of dominance megalithic tombs and passage graves of the Atlantic Megalithic culture appeared, especially in the Netherlands.  There have been at least five ice ages in the Earth's history which resulted in the presence or expansion of continental ice sheets. We are in an ice age today, called the Pleistocene or Quaternary glaciation, with the continent of Antartica and the island of Greenland covered by glaciers. This ice age began over 2.5 million years ago.
There have been at least five ice ages in the Earth's history which resulted in the presence or expansion of continental ice sheets. We are in an ice age today, called the Pleistocene or Quaternary glaciation, with the continent of Antartica and the island of Greenland covered by glaciers. This ice age began over 2.5 million years ago.  The chief successors to the Linear Ware culture were the Lengyel and Stroke-Ornamented, both in the east, and the Rossen, in the west. The Rossen covered much of Germany, the Low Countries, northeastern France, northern Switzerland and parts of Austria. They continued some of the features of the previous culture, including enclosed settlements and long houses, and practiced a mixed agriculture.
The chief successors to the Linear Ware culture were the Lengyel and Stroke-Ornamented, both in the east, and the Rossen, in the west. The Rossen covered much of Germany, the Low Countries, northeastern France, northern Switzerland and parts of Austria. They continued some of the features of the previous culture, including enclosed settlements and long houses, and practiced a mixed agriculture.




